- Introduction: Laktaattidehydrogenaasireaktio
- Malaatti-aspartaattisukkula
- Konventionaalinen (an)aerobinen glykolyysi
- Laktaattisukkulakonsepti
- LDH:n solunsisäinen kompartmentalisaatio: Fosfokreatiinisukkulan opit
- Laktaattimetabolian solunsisäinen lokeroituminen: Astrosyytti-neuronien laktaattisukkula
- Sytosolisen NAD+:n regenerointi
- Protonien sukkulointi ja mitokondriaalinen substraattikuljetus
- Metodologiset näkökohdat
- Yhteenveto
- Erittely eturistiriidoista
- Kiitokset
Introduction: Laktaattidehydrogenaasireaktio
Pyruvaatin pelkistymistä laktaatiksi, jota laktaattidehydrogenaasi (LDH; Pyruvate + NADH + H+ ⇌ Laktaatti + NAD+) katalysoi monien solujen sytosolissa, on pidetty aineenvaihdunnallisena ”umpikujaan” johtavana reaktiona (”umpikujaan”) (”umpikujaan”) (Luft, 2001; Quistorff ja Grunnet, 2011a), tai ehkä osuvammin metabolisena umpikujana (Barros, 2013), koska laktaatti voi liittyä uudelleen metaboliseen verkostoon vain pyruvaatin kautta. Nisäkkäillä LDH-reaktiota pidetään myös ”lähes tasapainoisena” (Spriet ym., 2000; Quistorff ja Grunnet, 2011a,b), mikä tarkoittaa, että reaktiota säätelevät lähinnä sen reaktanttien ja tuotteiden pitoisuudet eikä niinkään kehittyneemmät allosterisen säätelyn tai kovalenttisen modifioinnin keinot (Crabtree ja Newsholme, 1978). Koska LDH-reaktion tasapaino on kaukana oikealla (eli laktaatin muodostuminen suosii) (Williamson ym., 1967) LDH:n isomuodosta riippumatta (Quistorff ja Grunnet, 2011a,b), tästä saattaa seurata, että LDH harvoin suosii käänteisreaktiota (eli laktaatin hapettumista) in vivo. Huomattavaan laktaattihapettumiseen tarvittavan massavaikutussuhteen (/) pitäisi nimittäin ylittää LDH:n tasapainovakio. Kokeellinen todistusaineisto kuitenkin kumoaa yhä useammin käsityksen siitä, että LDH toimii yksisuuntaisesti in vivo, ja tukee sitä, että laktaatti toimii tärkeänä aineenvaihdunnan polttoaineena monissa kudoksissa, myös luustossa (Brooks ym., 1991; Bergman ym, 1999; Donovan ja Pagliassotti, 2000) ja sydänlihaksessa (Gertz ym., 1988; Chatham ym., 2001), maksassa (Skilleter ja Kun, 1972; Kline ym., 1986) ja aivoissa (Schurr ym., 1988; Bouzier-Sore ym., 2006; Wyss ym., 2011; Funfschilling ym., 2012; katsaus Barros, 2013). Tämän näkökulman tarkoituksena on esittää malli, jossa laktaatti on keskeisessä asemassa energeettisen substraatin sukkuloinnissa sytosolin (glykolyysi) ja mitokondrioiden (oksidatiivinen fosforylaatio) välillä. Tällaisen konseptin osatekijät on osoitettu sydämessä (Safer ym., 1971) ja luurankolihaksessa (Schantz, 1986), ne on myöhemmin laajennettu laktaattisukkulanäkökulmaksi (Stainsby ja Brooks, 1990; Brooks ym., 1999), ja niitä on tarkasteltu kattavasti (Gladden, 2004), ja niitä on jälleen kommentoitu (Gladden, 2007). Konseptia tukevat erityisesti viimeaikaiset tutkimukset hermosoluissa (Gellerich ym., 2012, 2013; Rueda ym., 2014). Vaikka tässä näkökulmassa hahmoteltu käsite ei sinänsä ole uusi (Safer et al., 1971), ilmeinen konventionaalisen tunnustuksen tai sen teoreettisten perusteiden hyväksynnän puute vaatii lisähuomiota.
Malaatti-aspartaattisukkula
Johtuen mitokondrioiden sisäisen kalvon läpäisemättömyydestä NAD+:lle ja NADH:lle, glykolyysin tuottama NADH aerobisissa olosuhteissa riippuu pelkistävien ekvivalenttien epäsuorasta siirrosta mitokondrioihin malaatti- aspartaattisukkulan (MAS) ja glyserolifosfaattisukkulan (glyserol-fosfaattisukkula) kautta. Näiden sukkuloiden uskotaan myös regeneroivan sytosolista NAD+:a, jota tarvitaan tukemaan glykolyyttistä virtausta NAD+:a vaativassa glyseraldehydi-3-fosfaattidehydrogenaasireaktiossa. MAS:n on osoitettu olevan pääasiallinen keino, jolla tämä tapahtuu useimmissa hapellisissa kudoksissa, ja se näyttää muodostavan pääasiallisen NADH-sukkulan kypsissä neuroneissa (Kauppinen ym., 1987; Ramos ym., 2003; Contreras ja Satrustegui, 2009; Gellerich ym., 2012). On myös hyvin todettu, että lisääntyneen soluenergiantarpeen ja/tai lisääntyneen glykolyyttisen virtauksen (esim. rasittavan liikunnan aikana) sekä hypoksian aikana laktaattipitoisuus nousee, koska LDH-reaktio helpottaa sytosolisen NAD+:n lisääntynyttä regeneroitumista (Robergs ym., 2004). Aivoissa laktaattipitoisuuden lisääntyminen verenkierrossa (esim. kuten liikunnan aikana) johtaa kuitenkin laktaatin poistumisen lisääntymiseen aivoissa (Quistorff ym., 2008; van Hall ym., 2009; Boumezbeur ym., 2010; Dienel, 2012). On myös ehdotettu, että lisääntynyt LDH-aktiivisuus (ja sitä kautta laktaatin tuotanto) yksinkertaisesti kompensoi MAS:n kyvyttömyyttä pysyä sytosolisen NAD+:n kysynnän tahdissa (Schantz, 1986). Neuroneissa MAS:n ja TCA-syklin Ca+2-aktivaatio on kilpailevaa siten, että alhaisemmat Ca+2-pitoisuudet stimuloivat MAS-aktiivisuutta aktivoimalla glutamaatti/aspartaattikantajaa (Contreras ja Satrustegui, 2009), kun taas korkeammat Ca+2-pitoisuudet aktivoivat α-ketoglutaraattidehydrogenaasia mitokondriaalisessa matriksissa, mikä rajoittaa MAS:n käytettävissä olevan α-ketoglutaraatin määrää (Contreras ja Satrustegui, 2009). On myös mahdollista, että laktaattia muodostuu jatkuvasti sytosolissa metabolisesta tilasta riippumatta ja että mitokondrioissa hapetettu laktaatti kytkeytyy MAS:ään. Esimerkiksi eristetyissä sydämen mitokondrioissa MAS:lla on ylikapasiteettia, mikä viittaa siihen, että MAS-aktiivisuus yksinään riittää ylläpitämään sytosolista NAD+ -regeneraatiota (Digerness ja Reddy, 1976). Miksi sitten levossa ja täysin aerobisissa olosuhteissa glykolyysin aikana syntyisi laktaattia, jos kaiken pyruvaatin pitäisi mennä mitokondrioihin oksidatiivista fosforylaatiota varten ja MAS:n pitäisi regeneroida riittävästi NAD+:a?
Konventionaalinen (an)aerobinen glykolyysi
Laktaatin ilmaantuminen ja katoaminen vaihtelevien aineenvaihduntatilojen aikana on ollut historian aikana monien arvailujen, ristiriitaisuuksien ja juonittelun aiheena. Laktaattiaineenvaihduntaa tutkivasta kirjallisuudesta on tehty monia katsauksia, joihin lukijat voidaan ohjata. Uusimpia ovat muun muassa (Cruz et al., 2012; Dienel, 2012; Kitaoka et al., 2012; Doherty ja Cleveland, 2013; Newington et al., 2013; Brooks, 2014; Schurr, 2014; Todd, 2014). Valitettavasti monet nykyaikaiset oppikirjat käyttävät edelleen pyruvaatin metabolista kohtaloa erottaakseen toisistaan kaksi glykolyysityyppiä: aerobinen (eli happea vaativa) ja anaerobinen (eli ilman happea tapahtuva). Hapen läsnä ollessa on sanottu, että pyruvaatti siirtyy mitokondrioihin kohtaamaan metabolisen loppunsa oksidatiivisen fosforylaation kautta, jonka nettotuloksena on mitokondrioiden ATP:n uudelleensynteesi ja hapenkulutus (eli hengitys) (Voet et al., 2011). Päinvastoin, kun happi on rajallinen, LDH pelkistää pyruvaatin sytosolissa laktaatiksi hapettaen samalla kofaktoriaan NADH:ta (Voet et al., 2011). Ongelmana tässä perinteisessä konstruktiossa on se, että se ei sovi hyvin yhteen joidenkin toistuvien tieteellisten havaintojen kanssa. On esimerkiksi hyvin tiedossa, että laktaattia tuotetaan ja kulutetaan täysin aerobisissa olosuhteissa. Itse asiassa terveillä, normoksisilla henkilöillä, jotka ovat levossa postabsorptiotilassa, voidaan odottaa, että pelkästään aivoista vapautuu noin 50 μmol-min-1 laktaattia (van Hall et al., 2009; van Hall, 2010). On selvää, että laktaatti on enemmän kuin anaerobisen glykolyysin umpikujaan päätyvä jätemetaboliitti; pikemminkin laktaatin sukkulointi koko elimistössä tarjoaa hyödyllisen näkökulman, jonka avulla voidaan tulkita kokeellista havaintoa.
Laktaattisukkulakonsepti
Kahta laktaattisukkulakonseptia on esitetty, jotka kuvaavat laktaatin liikkumista ja hyödyntämistä solujen sisällä ja solujen välillä (Brooks, 1998). Solunsisäisen laktaattisukkulan hypoteesin mukaan glykolyysin aikana muodostuvaa laktaattia voidaan käyttää jatkuvasti energianlähteenä saman solun sisällä (Brooks, 1998). Solujen väliseen tai solun ja solun väliseen laktaattisukkulaan kuuluu, että laktaattia syntyy ja viedään solusta toisen solun käyttöön (Brooks, 1998). Solun ja solun välinen laktaattisukkula on saavuttanut yleisen hyväksynnän; solunsisäisen laktaattisukkulan tarkempia yksityiskohtia tutkitaan kuitenkin edelleen. Hiljattain osoitimme sekä LDH:n fyysisen että toiminnallisen assosiaation mitokondrioiden kanssa luurankolihaksessa (Elustondo et al., 2013). Käyttämällä laserkeilaavaa konfokaalimikroskopiaa vahvistimme LDH:n kolokalisaation mitokondrioiden kalvoproteiinien kanssa rotan luurankolihaksessa. Havaitsimme, että rottien saponiinipermeabiloitujen luurankolihaskuitujen mitokondriot hapettivat laktaattia NAD+:n, malaatin ja ADP:n läsnä ollessa (Elustondo ym., 2013); toinen ryhmä havaitsi tämän samalla tavalla ihmisen kuiduissa (Jacobs ym., 2013). Tämän jälkeen pyruvaatti kuljetettiin mitokondrioihin, jossa pyruvaattidehydrogenaasi (PDH) hapetti sitä edelleen, sitten TCA-kierto, jolloin pelkistävät ekvivalentit stimuloivat hengitystä (Elustondo et al., 2013; Jacobs et al., 2013). Pystyimme estämään hengityksen vain 5μM alfa-syano-hydroksisinnamaatilla, joka on mitokondriaalisen pyruvaattikuljetuksen estäjä, mikä tukee entisestään sitä, että pyruvaattia, mutta ei laktaattia, kuljetetaan mitokondriomatriisiin. Nämä havainnot tukevat sitä, että LDH on strategisesti sijoitettu siten, että se on toiminnallisesti vuorovaikutuksessa mitokondrioiden kanssa, ja viittaavat siihen, että laktaatin hapettuminen tapahtuu lähellä mitokondrioiden sisäisen kalvon ulkopintaa. Miten solunsisäinen laktaattisukkula voisi toimia ehjässä solussa? Palataanpa MAS:iin.
In vivo sytosolista NAD+:a voisi teoriassa regeneroida sytosolista NAD+:a malaattidehydrogenaasilla mitokondriomatriisin ulkopuolella, osana MAS:ää. Kirjallisuus antaa jonkin verran tietoa eri kudoksista ja niiden mitokondrioiden sukkulointitoiminnasta. Aivoissa MAS:ää on pidetty tärkeimpänä sukkulointijärjestelmänä sytosolisen NADH:n saamiseksi mitokondrioihin (McKenna ym., 2006, ja viittaukset siihen); sitä vastoin glyseroli-fosfaattisukkulointi näyttää olevan vähemmän tärkeä (Nguyen ym., 2003). MAS:n ja välittäjäaine glutamaatin biosynteesin läheisestä yhteydestä on raportoitu (Palaiologos ym., 1988). Rotan aivoista eristetyistä mitokondrioista mitatut julkaistut MAS:n aktiivisuudet ovat luokkaa 26,7 nmol-min-1-mg-1 (Pardo ym., 2006). Synaptosomeissa MAS:n farmakologinen inhibitio johtaa laktaatin hapettumisen selvään (eli 50 %:n) vähenemiseen (McKenna et al., 1993), mikä tukee kuvassa 1 esitettyä mallia. Aivonsisäinen laktaatin tuotanto 13C-merkitystä glukoosista tukee edelleen käsitystä siitä, että laktaatti on tärkeä polttoaine neuroneille (Sampol et al., 2013).
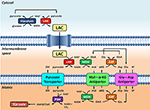
KUVIO 1. Laktaatin tuotanto 13C-merkitystä glukoosista. Kaavamainen esitys tässä Perspektiivissä hahmotellusta glykolyysin ja laktaattihapetuksen välisestä yhteydestä mitokondrioissa. Kaiken kaikkiaan glykolyysi tuottaa ATP:n lisäksi pyruvaattia ja NADH:ta. NAD+ voidaan palauttaa glykolyysiä varten pelkistämällä pyruviitti laktaatiksi (LAC) laktaattidehydrogenaasin (LDH) avulla. LAC voi diffundoitua mitokondrioihin, joissa LDH hapettaa sen pyruvaatiksi. NAD+ regeneroituu matriisin ulkopuolisen malaatti-aspartaattisukkulan malaatti-dehydrogenaasin (MDH) toimesta. Tämän jälkeen pyruvaatti kuljetetaan sisäisen mitokondriokalvon läpi matriisiin, jossa pyruvaattidehydrogenaasi (PDH) hapettaa sen asetyyli-CoA:ksi. Lyhenteet: α-KG, alfa-ketoglutaraatti; Glu, glutamaatti; AAT, aspartaattiaminotransferaasi; OAA, oksaloasetaatti; Mal, malaatti.
On huomattava, että MAS:llä voi olla rajansa. Suurilla sydämen työmäärillä on osoitettu, että sisemmän mitokondriokalvon α-ketoglutaraatti/malaattikuljettaja ei pysty kilpailemaan matriisin α-ketoglutaraattidehydrogenaasin kanssa niiden yhteisestä substraatista, α-ketoglutaraatista (O’Donnell ym., 2004). Tämä johtaa MAS:n rajoittavaan vaikutukseen ja sen NADH:n sukkulointiin mitokondrioihin. Tämän MAS:n rajoittamisen nettovaikutus olisi sytosolisten laktaattipitoisuuksien nousu, koska NAD+-regeneraatio LDH-reaktion kautta auttaisi säilyttämään homeostaattisen NAD+/NADH-arvon myös riittävän hapen läsnä ollessa. Tämä on todellakin klassinen ilmiö, joka havaitaan erityisen rasittavan liikunnan aikana, jolloin laktaattia voi kertyä vereen riittävästä hapen saatavuudesta huolimatta.
LDH:n solunsisäinen kompartmentalisaatio: Fosfokreatiinisukkulan opit
Ajatus yhdisteiden sukkuloinnista mitokondrioiden ja sytosolin välillä solun energiatarpeiden tyydyttämiseksi käyttäen lähes tasapainoentsyymejä ei varmasti ole uusi. Fosfokreatiinin (PCr) sukkuloinnissa on mukana erillisiä mitokondriaalisia ja sytosolista peräisin olevia kreatiinikinaasin (CK) isomuotoisia yhdisteitä, jotka siirtävät ”korkean energian” fosfaattia mitokondrioista sytosoliin. LDH-reaktion tavoin CK-reaktiota (fosfokreatiini + ADP + H+ ⇌ kreatiini + ATP) pidetään ”lähes tasapainoisena”, mikä suosii ATP:n resynteesiä. Kokeelliset todisteet osoittavat kuitenkin, että sydänlihassoluissa vain sytosolinen CK on itse asiassa tasapainossa tai lähellä sitä (Reviewed in Joubert et al., 2004). Mitokondriaalinen CK puolestaan, joka on lokalisoitu kalvojen väliseen tilaan, on siirtynyt tasapainosta, mikä suosii PCr:n netto-resynteesiä. Vastaavasti solunsisäiseen laktaattisukkulaan uskotaan osallistuvan kaksi erillistä LDH-populaatiota: sytosolinen ja mitokondriaalinen. Sytosolinen LDH olisi tasapainossa tai lähellä tasapainoa, kun taas mitokondriaalinen LDH olisi syrjäytynyt tasapainosta. Sytosolinen LDH suosisi laktaatin nettotuotantoa, kun taas mitokondriaalinen LDH suosisi laktaatin hapettumista. Ja aivan kuten adeniininukleotiditranslokaasi (ANT), joka kuljettaa ADP:tä matriisiin sisemmän mitokondriokalvon läpi ATP:n vastineeksi, helpottaa mitokondriaalisen CK:n syrjäytymistä tasapainosta kalvojen välisessä tilassa, niin myös pyruvaattikuljettaja kuljettaisi jatkuvasti pyruvaattia syrjäyttäen mitokondriaalisen LDH:n reaktion tasapainosta (kuva 1). Tällainen laktaattikuljetus hyötyisi LDH:n lokalisoitumisesta kalvojen väliseen tilaan lähelle sisempää mitokondriokalvoa, sitoutuneena ulomman mitokondriokalvon ulkopuolelle ulomman ja sisemmän mitokondriokalvon kosketuskohtiin tai molempiin. Jos solunsisäinen laktaattihapetus tapahtuu mitokondrioissa lokeroitumisen kautta, kuten PCr-sukkulan kohdalla, LDH:n solun sisäinen lokalisaatio mitokondrioissa, niiden kohdalla tai niiden ympärillä olisi merkittävä piirre.
Laktaattimetabolian solunsisäinen lokeroituminen: Astrosyytti-neuronien laktaattisukkula
Rikas ja kasvava joukko neuroenergisiä tutkimuksia tukee myös lokeroituneen laktaattimetabolian olemassaoloa vierekkäisten aivosolujen – nimittäin astrosyyttien ja neuronien – välillä (katsaus teoksessa Pellerin ja Magistretti, 2012). Solujen välisen laktaattisukkulan muunnos yleensä (Brooks, 2009), astrosyytti-neuroni-laktaattisukkula (Pellerin ja Magistretti, 2012) perustuu ajatukseen, että astrosyytit ovat pääasiassa glykolyyttisiä, kun taas neuronit ovat oksidatiivisempia (Bouzier-Sore ja Pellerin, 2013 ja viittaukset siihen). Metabolisen mallinnuksen avulla väitettiin, että suurempi metabolinen virtaus PDH: n ja mitokondriaalisten NADH-sukkuloiden kautta neuroneissa verrattuna astrosyytteihin sanelee välttämättä astrosyyttien nettolaktaatin vapautumisen astrosyytteistä ja neuronien hapettumisen (Neves et ai., 2012), mikä tukee monia kokeellisia havaintoja (ks. Bouzier-Sore ja Pellerin, 2013). Kuten aiemmin mainittiin, MAS muodostaa tärkeimmän mitokondriaalisen NADH-sukkulan kypsissä neuroneissa (Kauppinen et ai., 1987; Ramos et ai., 2003; Contreras ja Satrustegui, 2009; Gellerich et ai., 2012). Näin ollen laktaattia kuluttavan neuronin tärkeä ominaisuus voi hyvinkin olla sen korkea MAS-aktiivisuus (Neves et ai., 2012). Onko laktaatti-malaatti-aspartaattisukkulan aika? Onko lisäteoreettista tukea tällaiselle mallille, jossa laktaatti toimii pelkistävänä ekvivalenttina?
Sytosolisen NAD+:n regenerointi
Laktaatin hapettuminen mitokondrionissa tekee aerobisen glykolyysin edelleen mielekkääksi sallimalla sytosolisen NAD+:n regeneroinnin sytosolisen LDH:n avulla. Todisteet viljellyissä soluissa viittaavat todellakin erittäin labiiliin laktaatti/pyruvaatti-suhteeseen, joka vaihtelee homeostaattisen NAD+/NADH:n säilyttämiseksi (Sun ym., 2012). Tämä olisi solun kannalta edullista useista syistä. Ensinnäkin se tarjoaisi välittömän keinon, jonka avulla NAD+ voidaan regeneroida paikallisesti (eli sytosolissa, jossa glykolyysi tapahtuu); laktaatin (molekyylipaino = 89,07 g/mol) suurempi suhteellinen diffuusiokyky verrattuna NAD+:aan (molekyylipaino = 663,43 g/mol) tarkoittaa, että laktaatti voi diffundoitua helposti solusta lisääntyneen glykolyyttisen virran olosuhteissa (esim. voimakas liikunta, hypoksinen stressi), kun se ohjataan myös mitokondrioita kohti. Vähentyneen soluenergiantarpeen aikana laktaatin jatkuvaa tuotantoa paljon alhaisemman glykolyyttisen virtauksen aikana käytettäisiin edelleen homeostaattisen NAD+/NADH:n ylläpitämiseen solun sisällä sekä solunsisäisen laktaatin sukkuloinnin jatkuvaan kytkemiseen MAS:iin.
Protonien sukkulointi ja mitokondriaalinen substraattikuljetus
Laktaatin hapettuminen mitokondrioissa on aerobisen glykolyysin kannalta järkevää, koska laktaatin tuotanto sytosolissa kuluttaa tehokkaasti protonin (Robergs ym, 2004), minkä uskotaan auttavan lieventämään metabolista asidoosia, joka liittyy lisääntyneeseen ATP:n liikevaihtoon ja korkeisiin glykolyysinopeuksiin (Robergs et al., 2004). Laktaatin sytosolipitoisuus ylittää tyypillisesti pyruvaatin pitoisuuden vähintään kymmenkertaisesti, mikä tarkoittaa, että laktaatti eikä pyruviitti on vallitseva monokarboksylaatti, joka pääsee mitokondrioiden kalvojen väliseen tilaan (Brooks ym., 1999). Hapettamalla laktaattia mitokondrioiden kalvojen välisessä tilassa vapautuisi protoneja, joissa ne voisivat vaikuttaa mitokondrioiden protonien käyttövoiman ΔpH-komponenttiin sisemmän kalvon läpi (Santo-Domingo ja Demaurex, 2012) ja/tai kuljettaa niitä epäsuorasti mitokondrioihin MAS:n avulla. Kuten epäorgaanisen fosfaatin ja joidenkin muiden substraattien ja ionien kuljetuksen kohdalla (Santo-Domingo ja Demaurex, 2012), myös rotan maksan mitokondrioiden mitokondrioiden sisäisen kalvon läpi tapahtuva kantoaineen välityksellä tapahtuva pyruvaatin kuljetus näyttää olevan suoraan kytköksissä protonisymporttiin (tai OH-antiporttiin) (Papa et al., 1971; Halestrap, 1975). Laktaatin hapettuminen lähellä sisäisen mitokondriokalvon ulkopintaa, joka vapauttaa protonin, vaikuttaisi osaltaan ΔpH:hon ja sitä kautta pyruvaatin kuljetukseen matriisiin. Mitokondrioiden sisemmän kalvon vieressä LDH:n massavaikutussuhdetta (eli tuotteiden pitoisuuksia/reaktioiden pitoisuuksia) voisi suurelta osin helpottaa pyruvaatin ”vuotaminen”, kun sitä kuljetetaan jatkuvasti laktaattia läpäisemättömän mitokondrioiden sisemmän kalvon läpi, sekä NAD+:n runsas regeneroituminen MAS:n matriisin ulkopuolisen malaattidehydrogenaasin toimesta. Tässä mallissa pyruvaatin kuljetus mitokondrioiden sisemmän kalvon läpi vaikuttaisi suoraan laktaatin hapettumisnopeuteen matriisin ulkopuolella. Laktaatin hapettumista mitokondrioissa oletettaisiin näin ollen säädeltävän epäsuorasti PDH-reaktiolla matriisissa. Tämä olisi edullista, koska toisin kuin LDH:ta, PDH:ta säädellään voimakkaasti allosteriasta ja kovalenttisesta modifikaatiosta johtuen. Kuten mainittiin, mallinnus ennustaa korkean PDH-aktiivisuuden sanelevan hermosolujen laktaatin kulutusta in vivo (Neves et ai., 2012); ja korkea PDH-aktiivisuus luonnehtii myös laktaattia kuluttavia hermosoluja viljelyssä (Halim et ai., 2010).
Metodologiset näkökohdat
Jos mitokondriaalinen laktaattihapetus liittyy toiminnallisesti MAS:n aktiivisuuteen, olisi tärkeää sisällyttää MAS:n komponentit in vitro -analyyseihin mitokondriaalisen laktaattihapetuksen in vitro -analyyseistä, kuten itse malaatti tai oksaloasetaatti. Malaatti on todennäköinen valinta, koska sitä käytetään rutiininomaisesti hengityksen stimuloimiseksi in vitro, jolloin se kuljetetaan matriisiin ja mitokondriaalinen malaattidehydrogenaasi hapettaa sen oksaloasetaatiksi. Tämä oksaloasetaatti voi sitten kondensoitua asetyylikoA:n kanssa, jota muodostuu esimerkiksi pyruvaatin lisäämisen yhteydessä. Glutamaatin sisällyttäminen malaatin lisäksi mahdollistaa MAS:n täyden toiminnan mitokondriohengityksen tasolla. Tärkeä, mutta joskus huomiotta jätetty näkökohta asianmukaisessa mitokondriaalisen laktaattihapetuksen arvioinnissa on NAD+:n sisällyttäminen LDH-reaktion edellyttämänä kofaktorina ja ADP:n sisällyttäminen fosfaattiakseptorina oksidatiivisen fosforylaation stimuloimiseksi (ts. tilassa 3 tapahtuva hengitys). Lisäksi MAS:n matriisin ulkopuolinen komponentti sisältää malaattidehydrogenaasireaktion: oksaloasetaatti + NADH + H+ ⇌ malaatti + NAD+. Kokeelliset protokollat, joissa tutkitaan hengityshapenkulutusta lihaksesta eristetyissä mitokondrioissa käyttäen korkeita malaattipitoisuuksia (esim. 4 mM; Rasmussen ym., 2002), saattavat suosia malaattidehydrogenaasireaktiota käänteiseen suuntaan (eli malaatin hapettumista ja NADH + H+ -tuotantoa), kun sitä lisätään mitokondrioiden näytteeseen yhdessä NAD+:n kanssa. MAS:n palautuvuus on todellakin havaittu eristetyissä hepatosyyteissä (Berry, 1971) ja mitokondrioissa, joissa on rekonstruoitu MAS (Kunz ja Davis, 1991). Tämän MAS:n kääntymisen nettovaikutus hengitykseen olisi vähentää malaatin pääsyä mitokondrioihin ja muodostaa oksaloasetaattia. Vielä tärkeämpää on, että kääntäminen estäisi laktaatin hapettumisen pyruvaatiksi ja sitä seuraavan pyruvaatin kuljetuksen ja hapettumisen matriisissa. Ratkaisut näihin metodologisiin esteisiin, jotka haittaavat mitokondrioiden laktaattihapettumisen tarkkailua in vitro, edellyttävät ainakin yhden MAS-komponentin mukaan ottamista. Jos malaattia lisätään, sopiva pitoisuus olisi määritettävä kokeellisesti. ADP:n ja NAD+:n tai NADH:n (muistutetaan, että MAS tuottaa NAD+:a LDH-reaktiota varten) sisällyttäminen on myös välttämätöntä, jotta voidaan havaita tuntuvaa mitokondriaalista laktaattihapetusta.
Yhteenveto
Laktaatti-MAS on laktaatti- ja malaatti-aspartaattisukkuloiden vuorovaikutus pelkistävän tehon translokaatiossa mitokondrioihin erityisesti hapettuvissa, aineenvaihdunnallisesti aktiivisina pidetyissä soluissa.
Erittely eturistiriidoista
Tekijä ilmoittaa, että tutkimus tehtiin ilman kaupallisia tai taloudellisia suhteita, jotka voitaisiin tulkita mahdolliseksi eturistiriidaksi.
Kiitokset
Monet kiitokset tohtori Erich Gnaigerille (Innsbruckin lääketieteellinen yliopisto) ja tohtori George Brooksille (Kalifornian yliopisto, Berkeley), jotka kumpikin arvostelivat tämän työn aiempia luonnoksia.
Barros, L. F. (2013). Laktaatin metabolinen signalointi aivoissa. Trends Neurosci. 36, 396-404. doi: 10.1016/j.tins.2013.04.002
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Bergman, B. C., Wolfel, E. E., Butterfield, G. E., Lopaschuk, G. D., Casazza, G. A., Horning, M. A., et al. (1999). Aktiivisen lihaksen ja koko kehon laktaattikinetiikka kestävyysharjoittelun jälkeen miehillä. J. Appl. Physiol. (1985) 87, 1684-1696.
Pubmed Abstract | Pubmed Full Text | Google Scholar
Berry, M. N. (1971). Pyruvaatin energiariippuvainen pelkistyminen laktaatiksi rotan maksan ehjillä eristetyillä parenkyymisoluilla. Biokemia. Biophys. Res. Commun. 44, 1449-1456. doi: 10.1016/S0006-291X(71)80248-6
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Boumezbeur, F., Petersen, K. F., Cline, G. W., Mason, G. F., Behar, K. L., Shulman, G. I. ym. et al. (2010). Veren laktaatin osuus aivojen energia-aineenvaihduntaan ihmisillä mitattuna dynaamisella 13C-ydinmagneettiresonanssispektroskopialla. J. Neurosci. 30, 13983-13991. doi: 10.1523/JNEUROSCI.2040-10.2010
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Bouzier-Sore, A. K., and Pellerin, L. (2013). Astrosyyttien monimutkaisen metabolisen luonteen purkaminen. Front. Cell. Neurosci. 7:179. doi: 10.3389/fncel.2013.00179
Pubmed Abstract | Pubmed Full Text | Ref Full Text | CrossRef Full Text | Google Scholar
Bouzier-Sore, A. K., Voisin, P., Bouchaud, V., Bezancon, E., Franconi, J. M., ja Pellerin, L. (2006). Glukoosin ja laktaatin kilpailu hapettuvan energian substraatteina sekä neuroneissa että astrosyyteissä: vertaileva NMR-tutkimus. Eur. J. Neurosci. 24, 1687-1694. doi: 10.1111/j.1460-9568.2006.05056.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Brooks, G. A. (1998). Nisäkkäiden polttoaineen käyttö kestävän liikunnan aikana. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 120, 89-107. doi: 10.1016/S0305-0491(98)00025-X
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Brooks, G. A. (2009). Solu-solu- ja solunsisäiset laktaattisukkulat. J. Physiol. 587, 5591-5600. doi: 10.1113/jphysiol.2009.178350
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Brooks, G. A. (2014). ”Metaboliset järjestelmät: laktaatin muodostuminen ja hyödyntäminen”, in History of Exercise Physiology. Human Kinetics, ed C. M. Tipton (Champaign, IL: Human Kinetics), 447-475.
Google Scholar
Brooks, G. A., Butterfield, G. E., Wolfe, R. R., Groves, B. M., Mazzeo, R. S., Sutton, J. R., et al. (1991). Vähentynyt riippuvuus laktaatista harjoituksen aikana akklimatisoitumisen jälkeen 4300 metriin. J. Appl. Physiol. (1985). 71, 333-341.
Pubmed Abstract | Pubmed Full Text | Google Scholar
Brooks, G. A., Dubouchaud, H., Brown, M., Sicurello, J. P., and Butz, C. E. (1999). Mitokondriaalisen laktaattidehydrogenaasin ja laktaattihapetuksen rooli solunsisäisessä laktaattisukkulassa. Proc. Natl. Acad. Sci. U.S.A. 96, 1129-1134. doi: 10.1073/pnas.96.3.1129
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Chatham, J. C., Des Rosiers, C., ja Forder, J. R. (2001). Todisteet erillisistä reiteistä laktaatin imeytymiselle ja vapautumiselle perfusoidussa rotan sydämessä. Am. J. Physiol. Endocrinol. Metab. 281, E794-E802.
Pubmed Abstract | Pubmed Full Text | Google Scholar
Contreras, L., and Satrustegui, J. (2009). Kalsium-signalointi aivojen mitokondrioissa: malaatti-aspartaatti-NADH-sukkulan ja kalsium-uniporterin/mitokondriaalisen dehydrogenaasin reittien vuorovaikutus. J. Biol. Chem. 284, 7091-7099. doi: 10.1074/jbc.M808066200
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Crabtree, B., ja Newsholme, E. A. (1978). Metabolisen reitin lähes tasapainoreaktion herkkyys substraattipitoisuuden muutoksille. Eur. J. Biochem. 89, 19-22. doi: 10.1111/j.1432-1033.1978.tb20891.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Cruz, R. S., de Aguiar, R. A., Turnes, T., Penteado Dos Santos, R., de Oliveira, M. F., ja Caputo, F. (2012). Solunsisäinen sukkula: laktaatti-aerobinen aineenvaihdunta. ScientificWorldJournal 2012:420984. doi: 10.1100/2012/420984
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Dienel, G. A. (2012). Aivojen laktaattiaineenvaihdunta: löydöt ja kiistat. J. Cereb. Blood Flow Metab. 32, 1107-1138. doi: 10.1038/jcbfm.2011.175
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Digerness, S. B., and Reddy, W. J. (1976). Malaatti-aspartaattisukkula sydämen mitokondrioissa. J. Mol. Cell. Cardiol. 8, 779-785. doi: 10.1016/0022-2828(76)90084-5
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Doherty, J. R., and Cleveland, J. L. (2013). Laktaattiaineenvaihdunnan kohdistaminen syöpähoitoihin. J. Clin. Invest. 123, 3685-3692. doi: 10.1172/JCI69741
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Donovan, C. M., and Pagliassotti, M. J. (2000). Laktaatin hävittämisreittien kvantitatiivinen arviointi luustolihaksen kuitutyypeissä. Med. Sci. Sports Exerc. 32, 772-777. doi: 10.1097/00005768-200004000-00009
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Elustondo, P. A., White, A. E., Hughes, M. E., Brebner, K., Pavlov, E., and Kane, D. A. (2013). Laktaattidehydrogenaasin (LDH) fysikaalinen ja toiminnallinen yhteys luurankolihaksen mitokondrioiden kanssa. J. Biol. Chem. 288, 25309-25317. doi: 10.1074/jbc.M113.476648
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Funfschilling, U., Supplie, L. M., Mahad, D., Boretius, S., Saab, A. S., Edgar, J., et al. (2012). Glykolyyttiset oligodendrosyytit ylläpitävät myeliiniä ja pitkäaikaista aksonien eheyttä. Nature 485, 517-521. doi: 10.1038/nature11007
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Gellerich, F. N., Gizatullina, Z., Gainutdinov, T., Muth, K., Seppet, E., Orynbayeva, Z., et al. (2013). Aivojen mitokondrioiden energisoinnin hallinta sytosolisen kalsiumin avulla: mitokondrioiden kaasupoljin. IUBMB Life 65, 180-190. doi: 10.1002/iub.1131
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Gellerich, F. N., Gizatullina, Z., Trumbekaite, S., Korzeniewski, B., Gaynutdinov, T., Seppet, E., ym. al. (2012). Sytosolinen Ca2+ säätelee eristettyjen aivojen mitokondrioiden energisointia muodostamalla pyruvattia malaatti-aspartaattisukkulan kautta. Biochem. J. 443, 747-755. doi: 10.1042/BJ20110765
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Gertz, E. W., Wisneski, J. A., Stanley, W. C. ja Neese, R. A. (1988). Sydänlihaksen substraatin käyttö liikunnan aikana ihmisillä. Kahden hiilen leimatut hiilihydraatti-isotooppikokeet. J. Clin. Invest. 82, 2017-2025. doi: 10.1172/JCI113822
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Gladden, L. B. (2004). Laktaattiaineenvaihdunta: uusi paradigma kolmannelle vuosituhannelle. J. Physiol. 558, 5-30. doi: 10.1113/jphysiol.2003.058701
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Gladden, L. B. (2007). Onko luurankolihaksessa solunsisäinen laktaattisukkula? J. Physiol. 582, 899. doi: 10.1113/jphysiol.2007.138487
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Halestrap, A. P. (1975). Mitokondriaalinen pyruvaattikantaja. Kinetiikka ja spesifisyys substraateille ja inhibiittoreille. Biokemia. J. 148, 85-96.
Pubmed Abstract | Pubmed Full Text | Google Scholar
Halim, N. D., McFate, T., Mohyeldin, A., Okagaki, P., Korotchkina, L. G., Patel, M. S., et al. (2010). Pyruvaattidehydrogenaasin fosforylaatiotilanne erottaa viljeltyjen rotan aivojen astrosyyttien ja neuronien metaboliset fenotyypit. Glia 58, 1168-1176. doi: 10.1002/glia.20996
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Jacobs, R. A., Meinild, A. K., Nordsborg, N. B., and Lundby, C. (2013). Laktaatin hapettuminen ihmisen luurankolihaksen mitokondrioissa. Am. J. Physiol. Endocrinol. Metab. 304, E686-E689. doi: 10.1152/ajpendo.00476.2012
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Joubert, F., Mateo, P., Gillet, P., Gillet, Gillet, Gillet, Gillet, Gillet, Gillet, Gillet, Gillet, Gillet, Gillet, Gillet, Gillet, Gillet, Gillet, Gillet, Gillet, Gillet, B., B., B., B., B., B., B., Gillet, Gillet, Gillet, B., B., B., Gillet, B., B., B., B., B., J. CK-virtaus vai suora ATP-siirto: energiansiirtoreittien monipuolisuus osoitettuna NMR:llä perfusoidussa sydämessä. Mol. Cell. Biochem. 256-257, 43-58. doi: 10.1023/B:MCBI.0000009858.41434.fc
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kauppinen, R. A., Sihra, T. S., and Nicholls, D. G. (1987). Aminooksietikkahappo estää malaatti-aspartaattisukkulaa eristetyissä hermopäätteissä ja estää mitokondrioita käyttämästä glykolyyttisiä substraatteja. Biochim. Biophys. Acta 930, 173-178. doi: 10.1016/0167-4889(87)90029-2
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kitaoka, Y., Hoshino, D., and Hatta, H. (2012). Monokarboksylaattikuljettaja ja laktaattiaineenvaihdunta. J. Phys. Fitness Sports Med. 1, 247-252. doi: 10.7600/jpfsm.1.247
CrossRef Full Text | Google Scholar
Kline, E. S., Brandt, R. B., Laux, J. E., Spainhour, S. E., Higgins, E. S., Rogers, K. S., et al. (1986). L-laktaattidehydrogenaasin lokalisaatio mitokondrioissa. Arch. Biochem. Biophys. 246, 673-680. doi: 10.1016/0003-9861(86)90323-1
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kunz, W. S., and Davis, E. J. (1991). Pelkistävän potentiaalin palautuvan solunsisäisen siirron hallinta. Arch. Biochem. Biophys. 284, 40-46. doi: 10.1016/0003-9861(91)90260-P
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Luft, F. C. (2001). Maitohappoasidoosin päivitys tehohoidon kliinikoille. J. Am. Soc. Nephrol. 12 Suppl 17: S15-S19.
Pubmed Abstract | Pubmed Full Text | Google Scholar
McKenna, M. C., Tildon, J. T., Stevenson, J. H., Boatright, R. ja Huang, S. (1993). Energia-aineenvaihdunnan säätely synaptisissa päätepisteissä ja viljellyissä rotan aivojen astrosyyteissä: erot paljastuvat aminooksiasetaatin avulla. Dev. Neurosci. 15, 320-329. doi: 10.1159/000111351
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
McKenna, M. C., Waagepetersen, H. S., Schousboe, A. ja Sonnewald, U. (2006). Neuronaaliset ja astrosyyttiset sukkulamekanismit pelkistävien ekvivalenttien sytosoli-mitokondriosiirtoa varten: nykyiset todisteet ja farmakologiset välineet. Biochem. Pharmacol. 71, 399-407. doi: 10.1016/j.bcp.2005.10.011
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Neves, A., Costalat, R., and Pellerin, L. (2012). Aivosolujen metabolisten fenotyyppien ja energiasubstraattien käytön determinantit selvitetty mallinnuslähestymistavalla. PLoS Comput. Biol. 8:e1002686. doi: 10.1371/journal.pcbi.1002686
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Newington, J. T., Harris, R. A., and Cumming, R. C. (2013). Aineenvaihdunnan uudelleenarviointi Alzheimerin taudissa astrosyytti-neuroni laktaattisukkulamallin näkökulmasta. J. Neurodegener. Dis. 2013, 13. doi: 10.1155/2013/234572
CrossRef Full Text | Google Scholar
Nguyen, N. H., Brathe, A., ja Hassel, B. (2003). Glyserolin neuronaalinen otto ja metabolia sekä mitokondriaalisen glyseroli-3-fosfaattidehydrogenaasin neuronaalinen ilmentyminen. J. Neurochem. 85, 831-842. doi: 10.1046/j.1471-4159.2003.01762.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
O’Donnell, J. M., Kudej, R. K., LaNoue, K. F., Vatner, S. F. ja Lewandowski, E. D. (2004). Sytosolisen NADH:n rajallinen siirtyminen mitokondrioihin suuressa sydämen työkuormituksessa. Am. J. Physiol. Heart Circ. Physiol. 286, H2237-H2242. doi: 10.1152/ajpheart.01113.2003
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Palaiologos, G., Hertz, L., and Schousboe, A. (1988). Todisteet siitä, että aspartaattiaminotransferaasiaktiivisuus ja ketodikarboksylaattikantajan toiminta ovat välttämättömiä lähettimen glutamaatin biosynteesille. J. Neurochem. 51, 317-320. doi: 10.1111/j.1471-4159.1988.tb04872.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Papa, S., Francavilla, A., Paradies, G. ja Meduri, B. (1971). Pyruvaatin kuljetus rotan maksan mitokondrioissa. FEBS Lett. 12, 285-288. doi: 10.1016/0014-5793(71)80200-4
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Pardo, B., Contreras, L., Serrano, A., Ramos, M., Kobayashi, K., Iijima, M., et al. (2006). Aralarin olennainen rooli pienten Ca2+-signaalien välittämisessä neuronien mitokondrioihin. J. Biol. Chem. 281, 1039-1047. doi: 10.1074/jbc.M507270200
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Pellerin, L., and Magistretti, P. J. (2012). Makea kuusitoista ANLS:lle. J. Cereb. Blood Flow Metab. 32, 1152-1166. doi: 10.1038/jcbfm.2011.149
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Quistorff, B., ja Grunnet, N. (2011a). Korkea aivolaktaatti ei johdu laktaattidehydrogenaasi A/B -suhteen muutoksesta. Proc. Natl. Acad. Sci. U.S.A. 108, E21. author reply: E22. doi: 10.1073/pnas.1017750108
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Quistorff, B., ja Grunnet, N. (2011b). LDH:n isoentsyymimallilla ei ole fysiologista merkitystä; paitsi ehkä energia-aineenvaihdunnan nopeiden siirtymien aikana. Aging (Albany, NY) 3, 457-460.
Google Scholar
Quistorff, B., Secher, N. H., ja Van Lieshout, J. J. (2008). Laktaatti ruokkii ihmisen aivoja liikunnan aikana. FASEB J. 22, 3443-3449. doi: 10.1096/fj.08-106104
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Ramos, M., del Arco, A., Pardo, B., Martinez-Serrano, A., Martinez-Morales, J. R., Kobayashi, K., et al. (2003). Ca2+-säädellyn mitokondriaalisen aspartaatti-glutamaattikantajan aralar1:n kehitysmuutokset aivoissa ja näkyvä ilmentyminen selkäytimessä. Brain Res. Dev. Brain Res. 143, 33-46. doi: 10.1016/S0165-3806(03)00097-X
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Rasmussen, H. N., van Hall, G., ja Rasmussen, U. F. (2002). Laktaattidehydrogenaasi ei ole mitokondriaalinen entsyymi ihmisen ja hiiren vastus lateralis -lihaksessa. J. Physiol. 541, 575-580. doi: 10.1113/jphysiol.2002.019216
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Robergs, R. A., Ghiasvand, F., and Parker, D. (2004). Liikunnan aiheuttaman metabolisen asidoosin biokemia. Am. J. Physiol. Regul. Integr. Comp. Physiol. 287, R502-R516. doi: 10.1152/ajpregu.00114.2004
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Rueda, C. B., Llorente-Folch, I., Amigo, I., Contreras, L., Gonzalez-Sanchez, P., Martinez-Valero, P., et al. (2014). Mitokondrioiden toiminnan Ca2+-säätely neuroneissa. Biochim. Biophys. Acta 1837, 1617-1624. doi: 10.1016/j.bbabio.2014.04.010
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Safer, B., Smith, C. M., and Williamson, J. R. (1971). Pelkistävien ekvivalenttien kuljetuksen ohjaus mitokondriokalvon läpi perfusoidussa rotan sydämessä. J. Mol. Cell. Cardiol. 2, 111-124. doi: 10.1016/0022-2828(71)90065-4
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Sampol, D., Ostrofet, E., Jobin, M. L., Raffard, G., Sanchez, S., Bouchaud, V., et al. (2013). Glukoosi- ja laktaattiaineenvaihdunta hereillä olevalla ja stimuloidulla rotalla: (13)C-NMR-tutkimus. Front. Neuroenergetics 5:5. doi: 10.3389/fnene.2013.00005
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Santo-Domingo, J., and Demaurex, N. (2012). Näkökulmia: SGP-symposium mitokondrioiden fysiologiasta ja lääketieteestä: mitokondrioiden pH:n renessanssi. J. Gen. Physiol. 139, 415-423. doi: 10.1085/jgp.201110767
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Schantz, P. G. (1986). Ihmisen luurankolihaksen plastisuus ja erityisesti fyysisen harjoittelun vaikutukset NADH-sukkuloiden entsyymipitoisuuksiin sekä hitaiden ja nopeiden myofibrillaaristen proteiinien fenotyyppiseen ilmentymiseen. Acta Physiol. Scand. Suppl. 558, 1-62.
Pubmed Abstract | Pubmed Full Text | Google Scholar
Schurr, A. (2014). Aivojen glykolyysi: vuosisadan jatkuva väärinkäsitys ja väärinkäsitys. Front. Neurosci. 8:360. doi: 10.3389/fnins.2014.00360
CrossRef Full Text | Google Scholar
Schurr, A., West, C. A., and Rigor, B. M. (1988). Laktaatin tukema synaptinen toiminta rotan hippokampuksen viipalevalmisteessa. Science 240, 1326-1328. doi: 10.1126/science.3375817
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Skilleter, D. N., ja Kun, E. (1972). L-laktaatin hapettuminen maksan mitokondrioissa. Arch. Biochem. Biophys. 152, 92-104. doi: 10.1016/0003-9861(72)90197-X
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Spriet, L. L., Howlett, R. A., ja Heigenhauser, G. J. (2000). Entsymaattinen lähestymistapa ihmisen luurankolihaksen laktaattituotantoon harjoituksen aikana. Med. Sci. Sports Exerc. 32, 756-763. doi: 10.1097/00005768-200004000-00007
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Stainsby, W. N., and Brooks, G. A. (1990). Maitohappoaineenvaihdunnan säätely supistuvissa lihaksissa ja harjoituksen aikana. Exerc. Sport Sci. Rev. 18, 29-63. doi: 10.1249/00003677-199001000-00005
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Sun, F., Dai, C., Xie, J., and Hu, X. (2012). Biokemialliset kysymykset sytosolisen vapaan NAD/NADH-suhteen arvioinnissa. PLoS ONE 7:e34525. doi: 10.1371/journal.pone.0034525
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Todd, J. J. (2014). Laktaatti: arvokas fyysisen suorituskyvyn ja aivotoiminnan ylläpitämisen kannalta liikunnan aikana. Biosci. Horiz. 7, 1-7. doi: 10.1093/biohorizons/hzu001
CrossRef Full Text | Google Scholar
van Hall, G. (2010). Laktaattikinetiikka ihmisen kudoksissa levossa ja liikunnan aikana. Acta Physiol. (Oxf.) 199, 499-508. doi: 10.1111/j.1748-1716.2010.02122.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
van Hall, G., Stromstad, M., Rasmussen, P., Jans, O., Zaar, M., Gam, C., ym. et al. (2009). Veren laktaatti on tärkeä energianlähde ihmisen aivoille. J. Cereb. Blood Flow Metab. 29, 1121-1129. doi: 10.1038/jcbfm.2009.35
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Voet, D., Voet, J. G., and Pratt, C. W. (2011). Biokemian perusteet: Life at the Molecular Level. Hoboken, NJ: Wiley.
Google Scholar
Williamson, D. H., Lund, P., and Krebs, H. A. (1967). Vapaan nikotiiniamidi-adeniinidinukleotidin redox-tila rotan maksan sytoplasmassa ja mitokondrioissa. Biochem. J. 103, 514-527.
Pubmed Abstract | Pubmed Full Text | Google Scholar