- Úvod: Reakce laktátdehydrogenázy
- Malát-aspartátový člun
- Konvenční (an)aerobní glykolýza
- Koncept kyvadlového pohybu laktátu
- Intracelulární kompartmentalizace LDH: poučení z fosfokreatinového raketoplánu
- Mezibuněčná kompartmentalizace metabolismu laktátu: kyvadlová doprava laktátu mezi astrocyty a neurony
- Regenerace cytosolického NAD+
- Proton Shuttling and Mitochondrial Substrate Transport
- Metodologické úvahy
- Shrnutí
- Prohlášení o střetu zájmů
- Poděkování
Úvod: Reakce laktátdehydrogenázy
Redukce pyruvátu na laktát, katalyzovaná laktátdehydrogenázou (LDH; pyruvát + NADH + H+ ⇌ laktát + NAD+) v cytosolu mnoha buněk, byla považována za metabolickou „slepou uličku“ (Luft, 2001; Quistorff a Grunnet, 2011a), nebo možná výstižněji za metabolickou slepou uličku (Barros, 2013), protože laktát se může znovu zapojit do metabolické sítě pouze prostřednictvím pyruvátu. U savců je reakce LDH také považována za „téměř rovnovážnou“ (Spriet et al., 2000; Quistorff a Grunnet, 2011a,b), což znamená, že reakce je regulována především koncentracemi jejích reaktantů a produktů, nikoliv složitějšími prostředky alosterické regulace nebo kovalentní modifikace (Crabtree a Newsholme, 1978). Protože rovnováha reakce LDH leží daleko vpravo (tj. upřednostňuje se tvorba laktátu) (Williamson a kol., 1967) bez ohledu na izoformu LDH (Quistorff a Grunnet, 2011a,b), může z toho vyplývat, že LDH in vivo zřídka upřednostňuje opačnou reakci (tj. oxidaci laktátu). Hmotnostní akční poměr (/) potřebný pro znatelnou oxidaci laktátu by totiž musel překročit rovnovážnou konstantu pro LDH. Experimentální důkazy však stále více vyvracejí jakoukoli představu, že LDH působí in vivo jednosměrně, a podporují, že laktát slouží jako důležité metabolické palivo pro mnoho tkání, včetně kosterních (Brooks et al., 1991; Bergman et al., 1999; Donovan a Pagliassotti, 2000) a srdečního svalu (Gertz et al., 1988; Chatham et al., 2001), jater (Skilleter a Kun, 1972; Kline et al., 1986) a mozku (Schurr et al., 1988; Bouzier-Sore et al., 2006; Wyss et al., 2011; Funfschilling et al., 2012; přehled v Barros, 2013). Účelem této Perspektivy je předložit model, ve kterém je laktát ústředním prvkem při přesunu energetického substrátu mezi cytosolem (glykolýza) a mitochondriemi (oxidativní fosforylace). Součásti takové koncepce byly prokázány v srdci (Safer et al., 1971) a kosterním svalu (Schantz, 1986), později byly rozšířeny na perspektivu laktátového člunu (Stainsby a Brooks, 1990; Brooks et al., 1999) a komplexně přezkoumány (Gladden, 2004) a znovu komentovány (Gladden, 2007). Tento koncept podporuje zejména nedávný výzkum neuronálních buněk (Gellerich et al., 2012, 2013; Rueda et al., 2014). Ačkoli koncept nastíněný v této Perspektivě není sám o sobě nový (Safer et al., 1971), zjevný nedostatek konvenčního uznání nebo přijetí jeho teoretických základů si zaslouží další pozornost.
Malát-aspartátový člun
Vzhledem k nepropustnosti vnitřní mitochondriální membrány pro NAD+ a NADH závisí NADH generovaný glykolýzou za aerobních podmínek na nepřímém přenosu redukčních ekvivalentů do mitochondrií prostřednictvím malát-aspartátového člunu (MAS) a glycerol-fosfátového člunu. Předpokládá se, že tyto raketoplány také regenerují cytosolický NAD+ nezbytný pro podporu glykolytického toku při reakci glyceraldehyd-3-fosfát dehydrogenázy vyžadující NAD+. Bylo prokázáno, že MAS je převládajícím prostředkem, kterým k tomu dochází ve většině oxidativních tkání, a zdá se, že představuje hlavní kyvadlovou dopravu NADH ve zralých neuronech (Kauppinen et al., 1987; Ramos et al., 2003; Contreras a Satrustegui, 2009; Gellerich et al., 2012). Je také dobře známo, že za podmínek zvýšené potřeby buněčné energie a/nebo zvýšeného glykolytického toku (např. při namáhavém cvičení) a také při hypoxii se zvýší koncentrace laktátu, protože reakce LDH usnadňuje zvýšenou rychlost regenerace cytosolického NAD+ (Robergs et al., 2004). V mozku však zvýšení koncentrace laktátu v oběhu (např. jako při cvičení) vede ke zvýšení likvidace laktátu v mozku (Quistorff et al., 2008; van Hall et al., 2009; Boumezbeur et al., 2010; Dienel, 2012). Předpokládá se také, že zvýšená aktivita LDH (a následně produkce laktátu) jednoduše kompenzuje neschopnost MAS držet krok s poptávkou po cytosolickém NAD+ (Schantz, 1986). V neuronech je aktivace MAS a TCA cyklu kompetitivní, takže nižší hladiny Ca+2 stimulují aktivitu MAS aktivací glutamát/aspartátového přenašeče (Contreras a Satrustegui, 2009), zatímco vyšší koncentrace Ca+2 aktivují α-ketoglutarát dehydrogenázu v mitochondriální matrix, což omezuje množství α-ketoglutarátu dostupného pro MAS (Contreras a Satrustegui, 2009). Je také možné, že laktát se tvoří kontinuálně v cytosolu bez ohledu na metabolický stav a že laktát oxidovaný v mitochondriích je spojen s MAS. Například v izolovaných srdečních mitochondriích vykazuje MAS nadměrnou kapacitu, což naznačuje, že samotná aktivita MAS je dostatečná k udržení regenerace cytosolického NAD+ (Digerness a Reddy, 1976). Proč by tedy v klidu a za plně aerobních podmínek měl během glykolýzy vznikat laktát, když by veškerý pyruvát měl jít do mitochondrií k oxidativní fosforylaci a MAS by měla regenerovat dostatečné množství NAD+?
Konvenční (an)aerobní glykolýza
Vznik a zánik laktátu během různých metabolických stavů byl předmětem mnoha historických dohadů, sporů a intrik. Existuje mnoho přehledů literatury zkoumající metabolismus laktátu, na které lze čtenáře odkázat. Některé z novějších zahrnují (Cruz et al., 2012; Dienel, 2012; Kitaoka et al., 2012; Doherty a Cleveland, 2013; Newington et al., 2013; Brooks, 2014; Schurr, 2014; Todd, 2014). Bohužel mnoho současných učebnic stále používá metabolický osud pyruvátu k rozlišení dvou typů glykolýzy: aerobní (tj. vyžadující kyslík) a anaerobní (tj. bez kyslíku). V přítomnosti kyslíku prý pyruvát pokračuje do mitochondrií, kde se setká se svým metabolickým zánikem prostřednictvím oxidativní fosforylace, jejímž čistým výsledkem je mitochondriální resyntéza ATP a spotřeba kyslíku (tj. dýchání) (Voet et al., 2011). Naopak při nedostatku kyslíku je pyruvát v cytosolu redukován na laktát pomocí LDH, který přitom oxiduje svůj kofaktor NADH (Voet et al., 2011). Problémem této tradiční konstrukce je, že se dobře neslučuje s některými opakujícími se vědeckými pozorováními. Například je dobře známo, že laktát vzniká a spotřebovává se za plně aerobních podmínek. U zdravých normoxických jedinců v klidu v postabsorpčním stavu lze skutečně očekávat, že jen z mozku se uvolní přibližně 50 μmol-min-1 laktátu (van Hall et al., 2009; van Hall, 2010). Je zřejmé, že laktát není jen mrtvým odpadním metabolitem anaerobní glykolýzy, ale že kyvadlový pohyb laktátu v celém organismu poskytuje užitečnou perspektivu pro interpretaci experimentálních pozorování.
Koncept kyvadlového pohybu laktátu
Byly předloženy dva koncepty kyvadlového pohybu laktátu, které popisují pohyb a využití laktátu uvnitř buněk a mezi nimi (Brooks, 1998). Hypotéza intracelulárního laktátového člunu předpokládá, že laktát vznikající během glykolýzy může být neustále využíván jako zdroj energie v rámci téže buňky (Brooks, 1998). Mezibuněčný nebo mezibuněčný přenos laktátu zahrnuje laktát, který vzniká a je exportován z jedné buňky, aby byl přijat a využit jinou buňkou (Brooks, 1998). Laktátový člun mezi buňkami je všeobecně přijímán, avšak jemnější detaily intracelulárního laktátového člunu jsou stále předmětem zkoumání. Nedávno jsme prokázali jak fyzikální, tak funkční spojení LDH s mitochondriemi v kosterním svalu (Elustondo et al., 2013). Pomocí laserové skenovací konfokální mikroskopie jsme potvrdili kolokalizaci LDH s mitochondriálními membránovými proteiny v kosterním svalu potkana. Zjistili jsme, že mitochondrie v saponinem permeabilizovaných vláknech kosterního svalu potkanů oxidují laktát v přítomnosti NAD+, malátu a ADP (Elustondo et al., 2013); podobně to zjistila jiná skupina u lidských vláken (Jacobs et al., 2013). Pyruvát byl poté transportován do mitochondrií, kde byl dále oxidován pyruvátdehydrogenázou (PDH) a následně TCA cyklem, přičemž redukční ekvivalenty stimulovaly dýchání (Elustondo et al., 2013; Jacobs et al., 2013). Respiraci jsme byli schopni inhibovat pouhými 5 μM alfa-kyano-hydroxycinamátu, inhibitoru mitochondriálního transportu pyruvátu, což dále potvrzuje, že do mitochondriální matrix je transportován pyruvát, ale nikoliv laktát. Tato zjištění potvrzují, že LDH má strategickou polohu pro funkční interakci s mitochondriemi, a naznačují, že k oxidaci laktátu dochází v blízkosti vnějšího povrchu vnitřní mitochondriální membrány. Jak může intracelulární přenos laktátu fungovat v neporušené buňce? Vraťme se k MAS.
In vivo by teoreticky mohl být cytosolický NAD+ regenerován malátdehydrogenázou mimo mitochondriální matrix, jako součást MAS. Literatura poskytuje určitý přehled o různých tkáních a jejich mitochondriálních shuttlingových aktivitách. V mozku je MAS považována za nejdůležitější kyvadlový systém, kterým se dostává cytosolový NADH do mitochondrií (McKenna et al., 2006, a odkazy tamtéž); naopak glycerol-fosfátový kyvadlový systém se zdá být méně důležitý (Nguyen et al., 2003). Bylo totiž zaznamenáno těsné spojení MAS a biosyntézy neurotransmiteru glutamátu (Palaiologos et al., 1988). Publikované aktivity MAS měřené v mitochondriích izolovaných z mozku potkana se pohybují v řádu 26,7 nmol-min-1-mg-1 (Pardo et al., 2006). V synaptosomech vede farmakologická inhibice MAS k výraznému (tj. 50%) poklesu oxidace laktátu (McKenna et al., 1993), což podporuje model znázorněný na obrázku 1. Intracerebrální produkce laktátu z glukózy značené 13C dále podporuje představu, že laktát je důležitým palivem pro neurony (Sampol et al., 2013).
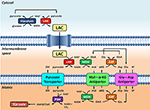
Obrázek 1. Schematické znázornění vazby mezi glykolýzou a oxidací laktátu v mitochondriích nastíněné v této Perspektivě. Celkově glykolýza poskytuje kromě ATP také pyruvát a NADH. NAD+ lze pro glykolýzu regenerovat redukcí pyruvátu na laktát (LAC) pomocí laktátdehydrogenázy (LDH). LAC může difundovat do mitochondrií, kde je LDH oxidován na pyruvát. NAD+ je regenerován extra-matrixovou malátdehydrogenázou (MDH) malát-aspartátového raketoplánu. Pyruvát je následně transportován přes vnitřní mitochondriální membránu do matrix, kde je poté oxidován pyruvátdehydrogenázou (PDH) na acetyl CoA. Zkratky: α-KG, alfa-ketoglutarát; Glu, glutamát; AAT, aspartát-aminotransferáza; OAA, oxaloacetát; Mal, malát.
Je třeba poznamenat, že MAS může mít své limity. Při vysokém srdečním zatížení bylo prokázáno, že α-ketoglutarát/malátový transportér vnitřní mitochondriální membrány nemůže soutěžit s matrixovou α-ketoglutarátdehydrogenázou o jejich společný substrát, α-ketoglutarát (O’Donnell et al., 2004). To má za následek limitující účinek na MAS a její přesun NADH do mitochondrií. Čistým účinkem tohoto omezení MAS by bylo zvýšení koncentrace cytosolového laktátu, protože regenerace NAD+ prostřednictvím reakce LDH by pomohla zachovat homeostatický NAD+/NADH, a to i v přítomnosti dostatečného množství kyslíku. To je ostatně klasický jev pozorovaný při zvláště namáhavém cvičení, kdy se laktát může hromadit v krvi navzdory dostatečné dostupnosti kyslíku.
Intracelulární kompartmentalizace LDH: poučení z fosfokreatinového raketoplánu
Představa o přesunu sloučenin mezi mitochondriemi a cytosolem za účelem uspokojení energetických požadavků buňky pomocí enzymů blízkých rovnováze jistě není nová. Do kyvadlové dopravy fosfokreatinu (PCr) jsou zapojeny různé izoformy mitochondriální a cytosolové kreatinkinázy (CK), které v podstatě přepravují „vysoce energetický“ fosfát z mitochondrií do cytosolu. Stejně jako reakce LDH je i reakce CK (fosfokreatin + ADP + H+ ⇌ kreatin + ATP) považována za „téměř rovnovážnou“ a upřednostňuje resyntézu ATP. Experimentální důkazy však ukazují, že v buňkách myokardu je ve skutečnosti v rovnováze nebo blízko rovnováhy pouze cytosolová CK (přehled v Joubert et al., 2004). Naproti tomu mitochondriální CK, lokalizovaná v mezimembránovém prostoru, je posunuta od rovnováhy a upřednostňuje čistou resyntézu PCr. Analogicky se předpokládá, že na intracelulárním přenosu laktátu se podílejí dvě odlišné populace LDH: cytosolová a mitochondriální. Cytosolová LDH by byla v rovnováze nebo blízko rovnováhy, zatímco mitochondriální LDH by byla z rovnováhy vytlačena. Cytosolová LDH by podporovala čistou produkci laktátu, zatímco mitochondriální LDH by podporovala oxidaci laktátu. A podobně jako adenin nukleotid translokáza (ANT), která transportuje ADP do matrix přes vnitřní mitochondriální membránu výměnou za ATP, usnadňuje vytěsnění z rovnováhy pro mitochondriální CK v mezimembránovém prostoru, tak i pyruvátový transportér by neustále transportoval pyruvát a vytěsňoval by z rovnováhy reakci mitochondriální LDH (obrázek 1). Takovému přenosu laktátu by prospěla lokalizace LDH v mezimembránovém prostoru v blízkosti vnitřní mitochondriální membrány, vazba na vnější stranu vnější mitochondriální membrány v místech kontaktu vnější a vnitřní mitochondriální membrány nebo obojí. Pokud by k intracelulární oxidaci laktátu mělo docházet v mitochondriích prostřednictvím kompartmentalizace, stejně jako v případě PCr shuttle, byla by buněčná lokalizace LDH v mitochondriích, u mitochondrií nebo v jejich okolí významným rysem.
Mezibuněčná kompartmentalizace metabolismu laktátu: kyvadlová doprava laktátu mezi astrocyty a neurony
Bohatý a stále rostoucí neuroenergetický výzkum rovněž podporuje existenci kompartmentalizovaného metabolismu laktátu mezi sousedními mozkovými buňkami – konkrétně astrocyty a neurony (přehled v Pellerin a Magistretti, 2012). Varianta mezibuněčného přenosu laktátu obecně (Brooks, 2009), přenos laktátu mezi astrocyty a neurony (Pellerin a Magistretti, 2012), je založena na myšlence, že astrocyty jsou převážně glykolytické, zatímco neurony jsou více oxidativní (Bouzier-Sore a Pellerin, 2013 a odkazy v nich). Pomocí přístupu metabolického modelování bylo argumentováno, že větší metabolický tok přes PDH a mitochondriální NADH shuttle v neuronech ve srovnání s astrocyty nutně diktuje čisté uvolňování laktátu astrocyty a oxidaci neurony (Neves et al., 2012), což podporuje mnoho experimentálních pozorování (přehled v Bouzier-Sore a Pellerin, 2013). Jak již bylo zmíněno, MAS představuje hlavní mitochondriální kyvadlovou dopravu NADH ve zralých neuronech (Kauppinen et al., 1987; Ramos et al., 2003; Contreras a Satrustegui, 2009; Gellerich et al., 2012). Proto může být důležitou vlastností neuronu spotřebovávajícího laktát jeho vysoká aktivita MAS (Neves et al., 2012). Přišel čas na laktát-malát-aspartátový shuttle? Existuje další teoretická podpora pro takový model, v němž laktát slouží jako redukční ekvivalent?
Regenerace cytosolického NAD+
Oxidace laktátu v mitochondrii dále dává smysl aerobní glykolýze tím, že umožňuje regeneraci cytosolického NAD+ pomocí cytosolické LDH. Důkazy v kultivovaných buňkách skutečně ukazují na vysoce labilní poměr laktát/pyruvát, který se mění, aby se zachoval homeostatický poměr NAD+/NADH (Sun et al., 2012). To by bylo pro buňku výhodné z několika důvodů. Zaprvé by to poskytlo okamžitý prostředek k lokální regeneraci NAD+ (tj. v cytosolu, kde probíhá glykolýza); větší relativní difuzibilita laktátu (molekulová hmotnost = 89,07 g/mol) oproti NAD+ (molekulová hmotnost = 663,43 g/mol) znamená, že laktát může snadno difundovat z buňky za podmínek zvýšeného glykolytického toku (např. intenzivní cvičení, hypoxický stres) a zároveň směřovat do mitochondrií. V době snížené potřeby buněčné energie by pokračující produkce laktátu při mnohem nižších rychlostech glykolytického toku byla stále využívána k udržení homeostatického NAD+/NADH v buňce, jakož i k pokračujícímu propojení intracelulárního přesunu laktátu s MAS.
Proton Shuttling and Mitochondrial Substrate Transport
Oxidace laktátu v mitochondriích dává smysl aerobní glykolýze, protože produkce laktátu v cytosolu účinně spotřebovává proton (Robergs et al., 2004), což pravděpodobně pomáhá zmírnit metabolickou acidózu spojenou se zvýšeným obratem ATP a vysokou rychlostí glykolýzy (Robergs et al., 2004). Cytosolická koncentrace laktátu obvykle nejméně desetkrát převyšuje koncentraci pyruvátu, což znamená, že laktát, a nikoli pyruvát, je převládajícím monokarboxylátem vstupujícím do mezimembránového prostoru mitochondrie (Brooks et al., 1999). Oxidací laktátu v mitochondriálním mezimembránovém prostoru by se uvolňovaly protony, které by mohly přispívat ke složce ΔpH mitochondriální protonové hybné síly přes vnitřní membránu (Santo-Domingo a Demaurex, 2012) a/nebo by byly nepřímo transportovány do mitochondrií prostřednictvím MAS. Stejně jako v případě transportu anorganického fosfátu a některých dalších substrátů a iontů (Santo-Domingo a Demaurex, 2012) se zdá, že transport pyruvátu přes vnitřní mitochondriální membránu v mitochondriích jater potkanů zprostředkovaný nosiči je přímo spojen s protonovým symportem (nebo OH-antiportem) (Papa et al., 1971; Halestrap, 1975). Oxidace laktátu v blízkosti vnějšího povrchu vnitřní mitochondriální membrány, při níž se uvolňuje proton, by přispívala k ΔpH a následně k transportu pyruvátu do matrix. V sousedství vnitřní mitochondriální membrány by mohl být hmotnostní akční poměr LDH (tj. koncentrace produktů/koncentrace reakcí) do značné míry usnadněn „vykrvácením“ pyruvátu, který je průběžně transportován přes vnitřní mitochondriální membránu nepropustnou pro laktát, a také velkorysou regenerací NAD+ pomocí extramatrixové malátdehydrogenázy MAS. V tomto modelu by transport pyruvátu přes vnitřní mitochondriální membránu přímo ovlivňoval rychlost oxidace laktátu mimo matrici. Očekávalo by se tedy, že oxidace laktátu v mitochondriích bude regulována nepřímo při reakci PDH v matrix. To by bylo výhodné, protože na rozdíl od LDH je PDH vysoce regulována prostřednictvím alosterie a kovalentní modifikace. Jak již bylo zmíněno, modelování předpovídá, že vysoká aktivita PDH diktuje spotřebu laktátu neurony in vivo (Neves et al., 2012); a vysoká aktivita PDH také charakterizuje neurony spotřebovávající laktát v kultuře (Halim et al.,
Metodologické úvahy
Pokud je mitochondriální oxidace laktátu funkčně spojena s aktivitou MAS, pak by bylo důležité zahrnout složky MAS do in vitro analýz mitochondriální oxidace laktátu, jako je samotný malát nebo oxaloacetát. Malát je pravděpodobnou volbou, protože je běžně součástí stimulace dýchání in vitro, kde je transportován do matrix a oxidován mitochondriální malátdehydrogenázou na oxaloacetát. Tento oxaloacetát pak může kondenzovat s acetyl coA, který vzniká například po přidání pyruvátu. Zahrnutí glutamátu vedle malátu umožňuje plnou činnost MAS na úrovni mitochondriálního dýchání. Důležitým, ale někdy přehlíženým aspektem vhodného posouzení mitochondriální oxidace laktátu je zahrnutí NAD+ jako nezbytného kofaktoru pro reakci LDH a ADP jako akceptoru fosfátu pro stimulaci oxidativní fosforylace (tj. dýchání ve stavu 3). Také mimomatriční složka MAS zahrnuje reakci malátdehydrogenázy: oxaloacetát + NADH + H+ ⇌ malát + NAD+. Experimentální protokoly zkoumající respirační spotřebu kyslíku v izolovaných mitochondriích ze svalu za použití vysokých koncentrací malátu (např. 4 mM; Rasmussen et al., 2002) mohou při přidání do mitochondriálního vzorku v kombinaci s NAD+ upřednostňovat reakci malátdehydrogenázy v opačném směru (tj. oxidaci malátu a produkci NADH + H+). Reverzibilita MAS byla skutečně pozorována v izolovaných hepatocytech (Berry, 1971) a mitochondriích s rekonstituovanou MAS (Kunz a Davis, 1991). Čistým účinkem této reverze MAS na dýchání by bylo snížení vstupu malátu do mitochondrií za vzniku oxaloacetátu. Ještě důležitější je, že obrácení by zabránilo oxidaci laktátu na pyruvát a následnému transportu a oxidaci pyruvátu v matrix. Řešení těchto metodických překážek pozorování mitochondriální oxidace laktátu in vitro zahrnuje zahrnutí alespoň jedné složky MAS. V případě přidání malátu by měla být vhodná koncentrace stanovena experimentálně. Zahrnutí ADP a NAD+ nebo NADH (připomeňme, že MAS vytvoří NAD+ pro reakci LDH) je rovněž nezbytné k pozorování znatelné mitochondriální oxidace laktátu.
Shrnutí
MAS laktátu je interakce mezi laktátem a malát-aspartátem, která přenáší redukční energii do mitochondrií, zejména v oxidativních, metabolicky aktivních buňkách.
Prohlášení o střetu zájmů
Autor prohlašuje, že výzkum byl prováděn bez jakýchkoli komerčních nebo finančních vztahů, které by mohly být chápány jako potenciální střet zájmů.
Poděkování
Velmi děkuji Dr. Erichu Gnaigerovi (Lékařská univerzita v Innsbrucku) a Dr. Georgi Brooksovi (Kalifornská univerzita v Berkeley) za každou z kritik dřívějších návrhů této práce.
Barros, L. F. (2013). Metabolická signalizace laktátem v mozku. Trends Neurosci. 36, 396-404. doi: 10.1016/j.tins.2013.04.002
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Bergman, B. C., Wolfel, E. E., Butterfield, G. E., Lopaschuk, G. D., Casazza, G. A., Horning, M. A., et al. (1999). Kinetika aktivních svalů a celotělového laktátu po vytrvalostním tréninku u mužů. J. Appl. Physiol. (1985) 87, 1684-1696.
Pubmed Abstract | Pubmed Full Text | Google Scholar
Berry, M. N. (1971). Energeticky závislá redukce pyruvátu na laktát intaktními izolovanými parenchymovými buňkami z jater potkana. Biochem. Biophys. Res. Commun. 44, 1449-1456. doi: 10.1016/S0006-291X(71)80248-6
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Boumezbeur, F., Petersen, K. F., Cline, G. W., Mason, G. F., Behar, K. L., Shulman, G. I., et al. (2010). The contribution of blood lactate to brain energy metabolism in humans measured by dynamic 13C nuclear magnetic resonance spectroscopy [Podíl krevního laktátu na energetickém metabolismu mozku u lidí měřený pomocí dynamické 13C nukleární magnetické rezonance]. J. Neurosci. 30, 13983-13991. doi: 10.1523/JNEUROSCI.2040-10.2010
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Bouzier-Sore, A. K., and Pellerin, L. (2013). Odhalování komplexní metabolické povahy astrocytů. Front. Cell. Neurosci. 7:179. doi: 10.3389/fncel.2013.00179
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Bouzier-Sore, A. K., Voisin, P., Bouchaud, V., Bezancon, E., Franconi, J. M., and Pellerin, L. (2006). Konkurence mezi glukózou a laktátem jako oxidačními energetickými substráty v neuronech i astrocytech: srovnávací NMR studie. Eur. J. Neurosci. 24, 1687-1694. doi: 10.1111/j.1460-9568.2006.05056.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Brooks, G. A. (1998). Využití paliv u savců během trvalé fyzické zátěže. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 120, 89-107. doi: 10.1016/S0305-0491(98)00025-X
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Brooks, G. A. (2009). Cell-cell and intracellular lactate shuttles (Přenosy laktátu mezi buňkami a uvnitř buněk). J. Physiol. 587, 5591-5600. doi: 10.1113/jphysiol.2009.178350
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Brooks, G. A. (2014). „Metabolické systémy: tvorba a využití laktátu,“ in History of Exercise Physiology. Human Kinetics, ed C. M. Tipton (Champaign, IL: Human Kinetics), 447-475.
Google Scholar
Brooks, G. A., Butterfield, G. E., Wolfe, R. R., Groves, B. M., Mazzeo, R. S., Sutton, J. R. a další (1991). Decreased reliance on lactate during exercise after acclimatization to 4 300 m. J. Appl. Physiol. (1985). 71, 333-341.
Pubmed Abstract | Pubmed Full Text | Google Scholar
Brooks, G. A., Dubouchaud, H., Brown, M., Sicurello, J. P., and Butz, C. E. (1999). Role mitochondriální laktátdehydrogenázy a oxidace laktátu v intracelulárním přenosu laktátu. Proc. Natl. Acad. Sci. U.S.A. 96, 1129-1134. doi: 10.1073/pnas.96.3.1129
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Chatham, J. C., Des Rosiers, C., and Forder, J. R. (2001). Důkaz oddělených cest pro příjem a uvolňování laktátu perfundovaným srdcem potkana. Am. J. Physiol. Endocrinol. Metab. 281, E794-E802.
Pubmed Abstract | Pubmed Full Text | Google Scholar
Contreras, L., and Satrustegui, J. (2009). Signalizace vápníku v mozkových mitochondriích: souhra cest malát-asparagin NADH shuttle a vápníkového uniporteru/mitochondriální dehydrogenázy. J. Biol. Chem. 284, 7091-7099. doi: 10.1074/jbc.M808066200
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Crabtree, B., and Newsholme, E. A. (1978). Citlivost téměř rovnovážné reakce v metabolické dráze na změny koncentrace substrátu. Eur. J. Biochem. 89, 19-22. doi: 10.1111/j.1432-1033.1978.tb20891.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Cruz, R. S., de Aguiar, R. A., Turnes, T., Penteado Dos Santos, R., de Oliveira, M. F., and Caputo, F. (2012). Intracelulární kyvadlová doprava: aerobní metabolismus laktátu. ScientificWorldJournal 2012:420984. doi: 10.1100/2012/420984
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Dienel, G. A. (2012). Metabolismus mozkového laktátu: objevy a kontroverze. J. Cereb. Blood Flow Metab. 32, 1107-1138. doi: 10.1038/jcbfm.2011.175
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Digerness, S. B., and Reddy, W. J. (1976). Malát-aspartátový člun v srdečních mitochondriích. J. Mol. Cell. Cardiol. 8, 779-785. doi: 10.1016/0022-2828(76)90084-5
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Doherty, J. R., and Cleveland, J. L. (2013). Cílení metabolismu laktátu pro léčbu rakoviny. J. Clin. Invest. 123, 3685-3692. doi: 10.1172/JCI69741
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Donovan, C. M., and Pagliassotti, M. J. (2000). Kvantitativní hodnocení cest pro likvidaci laktátu v typech vláken kosterního svalu. Med. Sci. Sports Exerc. 32, 772-777. doi: 10.1097/00005768-200004000-00009
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Elustondo, P. A., White, A. E., Hughes, M. E., Brebner, K., Pavlov, E., and Kane, D. A. (2013). Fyzikální a funkční spojení laktátdehydrogenázy (LDH) s mitochondriemi kosterního svalu. J. Biol. Chem. 288, 25309-25317. doi: 10.1074/jbc.M113.476648
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Funfschilling, U., Supplie, L. M., Mahad, D., Boretius, S., Saab, A. S., Edgar, J., et al. (2012). Glykolytické oligodendrocyty udržují myelin a dlouhodobou integritu axonů. Nature 485, 517-521. doi: 10.1038/nature11007
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Gellerich, F. N., Gizatullina, Z., Gainutdinov, T., Muth, K., Seppet, E., Orynbayeva, Z., et al. (2013). Řízení energetizace mozkových mitochondrií cytosolickým vápníkem: mitochondriální plynový pedál. IUBMB Life 65, 180-190. doi: 10.1002/iub.1131
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Gellerich, F. N., Gizatullina, Z., Trumbekaite, S., Korzeniewski, B., Gaynutdinov, T., Seppet, E., et al. (2012). Cytosolický Ca2+ reguluje energetizaci izolovaných mozkových mitochondrií tvorbou pyruvátu prostřednictvím malát-aspartátového člunu. Biochem. J. 443, 747-755. doi: 10.1042/BJ20110765
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Gertz, E. W., Wisneski, J. A., Stanley, W. C., and Neese, R. A. (1988). Myocardial substrate utilization during exercise in humans (Využití substrátů v myokardu během cvičení u lidí). Dual carbon-labeled carbohydrate isotope experiments. J. Clin. Invest. 82, 2017-2025. doi: 10.1172/JCI113822
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Gladden, L. B. (2004). Laktátový metabolismus: nové paradigma pro třetí tisíciletí. J. Physiol. 558, 5-30. doi: 10.1113/jphysiol.2003.058701
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Gladden, L. B. (2007). Existuje v kosterním svalu intracelulární kyvadlová doprava laktátu? J. Physiol. 582, 899. doi: 10.1113/jphysiol.2007.138487
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Halestrap, A. P. (1975). Mitochondriální pyruvátový přenašeč. Kinetika a specifita pro substráty a inhibitory. Biochem. J. 148, 85-96.
Pubmed Abstract | Pubmed Full Text | Google Scholar
Halim, N. D., McFate, T., Mohyeldin, A., Okagaki, P., Korotchkina, L. G., Patel, M. S., et al. (2010). Stav fosforylace pyruvátdehydrogenázy rozlišuje metabolické fenotypy kultivovaných potkaních mozkových astrocytů a neuronů. Glia 58, 1168-1176. doi: 10.1002/glia.20996
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Jacobs, R. A., Meinild, A. K., Nordsborg, N. B., and Lundby, C. (2013). Oxidace laktátu v mitochondriích lidského kosterního svalu. Am. J. Physiol. Endocrinol. Metab. 304, E686-E689. doi: 10.1152/ajpendo.00476.2012
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Joubert, F., Mateo, P., Gillet, B., Beloeil, J. C., Mazet, J. L., and Hoerter, J. A. (2004). Tok CK nebo přímý přenos ATP: všestrannost cest přenosu energie prokázaná pomocí NMR v perfundovaném srdci. Mol. Cell. Biochem. 256-257, 43-58. doi: 10.1023/B:MCBI.0000009858.41434.fc
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kauppinen, R. A., Sihra, T. S., and Nicholls, D. G. (1987). Aminooxyoctová kyselina inhibuje malát-aspartátový shuttle v izolovaných nervových terminálech a zabraňuje mitochondriím využívat glykolytické substráty. Biochim. Biophys. Acta 930, 173-178. doi: 10.1016/0167-4889(87)90029-2
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kitaoka, Y., Hoshino, D., and Hatta, H. (2012). Monokarboxylátový transportér a metabolismus laktátu. J. Phys. Fitness Sports Med. 1, 247-252. doi: 10.7600/jpfsm.1.247
CrossRef Full Text | Google Scholar
Kline, E. S., Brandt, R. B., Laux, J. E., Spainhour, S. E., Higgins, E. S., Rogers, K. S., et al. (1986). Lokalizace L-laktátdehydrogenázy v mitochondriích. Arch. Biochem. Biophys. 246, 673-680. doi: 10.1016/0003-9861(86)90323-1
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kunz, W. S., and Davis, E. J. (1991). Řízení reverzibilního vnitrobuněčného přenosu redukčního potenciálu. Arch. Biochem. Biophys. 284, 40-46. doi: 10.1016/0003-9861(91)90260-P
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Luft, F. C. (2001). Lactic acidosis update for critical care clinicians (Aktuální informace o laktátové acidóze pro lékaře v kritické péči). J. Am. Soc. Nephrol. 12 Suppl 17: S15-S19.
Pubmed Abstract | Pubmed Full Text | Google Scholar
McKenna, M. C., Tildon, J. T., Stevenson, J. H., Boatright, R., and Huang, S. (1993). Regulace energetického metabolismu v synaptických terminálech a kultivovaných astrocytech mozku potkana: rozdíly odhalené pomocí aminooxyacetátu. Dev. Neurosci. 15, 320-329. doi: 10.1159/000111351
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
McKenna, M. C., Waagepetersen, H. S., Schousboe, A., and Sonnewald, U. (2006). Neuronální a astrocytární shuttle mechanismy pro cytosolický-mitochondriální přenos redukčních ekvivalentů: současné důkazy a farmakologické nástroje. Biochem. Pharmacol. 71, 399-407. doi: 10.1016/j.bcp.2005.10.011
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Neves, A., Costalat, R., and Pellerin, L. (2012). Determinanty fenotypů metabolismu mozkových buněk a využití energetických substrátů odhalené pomocí modelového přístupu. PLoS Comput. Biol. 8:e1002686. doi: 10.1371/journal.pcbi.1002686
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Newington, J. T., Harris, R. A., and Cumming, R. C. (2013). Přehodnocení metabolismu u alzheimerovy choroby z pohledu modelu kyvadlové dopravy laktátu mezi astrocyty a neurony. J. Neurodegener. Dis. 2013, 13. doi: 10.1155/2013/234572
CrossRef Full Text | Google Scholar
Nguyen, N. H., Brathe, A., and Hassel, B. (2003). Neuronální příjem a metabolismus glycerolu a neuronální exprese mitochondriální glycerol-3-fosfát dehydrogenázy. J. Neurochem. 85, 831-842. doi: 10.1046/j.1471-4159.2003.01762.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
O’Donnell, J. M., Kudej, R. K., LaNoue, K. F., Vatner, S. F., and Lewandowski, E. D. (2004). Omezený přenos cytosolického NADH do mitochondrií při vysokém srdečním zatížení. Am. J. Physiol. Heart Circ. Physiol. 286, H2237-H2242. doi: 10.1152/ajpheart.01113.2003
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Palaiologos, G., Hertz, L., and Schousboe, A. (1988). Důkaz, že aktivita aspartátaminotransferázy a funkce ketodikarboxylátového přenašeče jsou nezbytné pro biosyntézu transmiteru glutamátu. J. Neurochem. 51, 317-320. doi: 10.1111/j.1471-4159.1988.tb04872.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Papa, S., Francavilla, A., Paradies, G., and Meduri, B. (1971). Transport pyruvátu v mitochondriích jater potkana. FEBS Lett. 12, 285-288. doi: 10.1016/0014-5793(71)80200-4
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Pardo, B., Contreras, L., Serrano, A., Ramos, M., Kobayashi, K., Iijima, M., et al. (2006). Zásadní úloha aralaru při přenosu malých signálů Ca2+ do neuronálních mitochondrií. J. Biol. Chem. 281, 1039-1047. doi: 10.1074/jbc.M507270200
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Pellerin, L., and Magistretti, P. J. (2012). Sladkých šestnáct pro ANLS. J. Cereb. Blood Flow Metab. 32, 1152-1166. doi: 10.1038/jcbfm.2011.149
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Quistorff, B., and Grunnet, N. (2011a). Vysoká hladina laktátu v mozku není způsobena posunem poměru laktátdehydrogenázy A/B. Proc. Natl. Acad. Sci. U.S.A. 108, E21. odpověď autora: E22. doi: 10.1073/pnas.1017750108
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Quistorff, B., and Grunnet, N. (2011b). Izoenzymový vzorec LDH nehraje fyziologickou roli; snad s výjimkou rychlých přechodů v energetickém metabolismu. Aging (Albany, NY) 3, 457-460.
Google Scholar
Quistorff, B., Secher, N. H., a Van Lieshout, J. J. (2008). Laktát pohání lidský mozek během cvičení. FASEB J. 22, 3443-3449. doi: 10.1096/fj.08-106104
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Ramos, M., del Arco, A., Pardo, B., Martinez-Serrano, A., Martinez-Morales, J. R., Kobayashi, K., et al. (2003). Vývojové změny Ca2+ regulovaného mitochondriálního aspartát-glutamátového přenašeče aralar1 v mozku a výrazná exprese v míše. Brain Res. Dev. Brain Res. 143, 33-46. doi: 10.1016/S0165-3806(03)00097-X
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Rasmussen, H. N., van Hall, G., and Rasmussen, U. F. (2002). Laktátdehydrogenáza není mitochondriální enzym v lidském a myším svalu vastus lateralis. J. Physiol. 541, 575-580. doi: 10.1113/jphysiol.2002.019216
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Robergs, R. A., Ghiasvand, F., and Parker, D. (2004). Biochemie metabolické acidózy vyvolané cvičením. Am. J. Physiol. Regul. Integr. Comp. Physiol. 287, R502-R516. doi: 10.1152/ajpregu.00114.2004
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Rueda, C. B., Llorente-Folch, I., Amigo, I., Contreras, L., Gonzalez-Sanchez, P., Martinez-Valero, P., et al. (2014). Regulace Ca2+ funkce mitochondrií v neuronech. Biochim. Biophys. Acta 1837, 1617-1624. doi: 10.1016/j.bbabio.2014.04.010
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Safer, B., Smith, C. M., and Williamson, J. R. (1971). Kontrola transportu redukčních ekvivalentů přes mitochondriální membránu v perfundovaném srdci potkana. J. Mol. Cell. Cardiol. 2, 111-124. doi: 10.1016/0022-2828(71)90065-4
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Sampol, D., Ostrofet, E., Jobin, M. L., Raffard, G., Sanchez, S., Bouchaud, V., et al. (2013). Metabolismus glukózy a laktátu u bdělého a stimulovaného potkana: studie (13)C-NMR. Front. Neuroenergetics 5:5. doi: 10.3389/fnene.2013.00005
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Santo-Domingo, J., and Demaurex, N. (2012). Perspektivy: SGP sympozium o mitochondriální fyziologii a medicíně: renesance mitochondriálního pH. J. Gen. Physiol. 139, 415-423. doi: 10.1085/jgp.201110767
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Schantz, P. G. (1986). Plasticita lidského kosterního svalu se zvláštním zřetelem k účinkům fyzického tréninku na hladiny enzymů NADH shuttle a fenotypovou expresi pomalých a rychlých myofibrilárních proteinů. Acta Physiol. Scand. Suppl. 558, 1-62.
Pubmed Abstract | Pubmed Full Text | Google Scholar
Schurr, A. (2014). Mozková glykolýza: století přetrvávajících nedorozumění a mylných představ. Front. Neurosci. 8:360. doi: 10.3389/fnins.2014.00360
CrossRef Full Text | Google Scholar
Schurr, A., West, C. A., and Rigor, B. M. (1988). Synaptická funkce podporovaná laktátem v preparátu hipokampálního plátku potkana. Science 240, 1326-1328. doi: 10.1126/science.3375817
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Skilleter, D. N., and Kun, E. (1972). Oxidace L-laktátu jaterními mitochondriemi. Arch. Biochem. Biophys. 152, 92-104. doi: 10.1016/0003-9861(72)90197-X
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Spriet, L. L., Howlett, R. A., and Heigenhauser, G. J. (2000). Enzymatický přístup k produkci laktátu v lidském kosterním svalu během cvičení. Med. Sci. Sports Exerc. 32, 756-763. doi: 10.1097/00005768-200004000-00007
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Stainsby, W. N., and Brooks, G. A. (1990). Řízení metabolismu kyseliny mléčné v kontrahujících se svalech a během cvičení. Exerc. Sport Sci. Rev. 18, 29-63. doi: 10.1249/00003677-199001000-00005
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Sun, F., Dai, C., Xie, J., and Hu, X. (2012). Biochemické problémy při odhadu poměru cytosolického volného NAD/NADH. PLoS ONE 7:e34525. doi: 10.1371/journal.pone.0034525
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Todd, J. J. (2014). Laktát: cenný pro fyzický výkon a udržení mozkových funkcí během cvičení. Biosci. Horiz. 7, 1-7. doi: 10.1093/biohorizons/hzu001
CrossRef Full Text | Google Scholar
van Hall, G. (2010). Kinetika laktátu v lidských tkáních v klidu a při cvičení. Acta Physiol. (Oxf.) 199, 499-508. doi: 10.1111/j.1748-1716.2010.02122.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
van Hall, G., Stromstad, M., Rasmussen, P., Jans, O., Zaar, M., Gam, C., et al. (2009). Krevní laktát je důležitým zdrojem energie pro lidský mozek. J. Cereb. Blood Flow Metab. 29, 1121-1129. doi: 10.1038/jcbfm.2009.35
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Voet, D., Voet, J. G., and Pratt, C. W. (2011). Základy biochemie: Základy biochemie: Život na molekulární úrovni. Hoboken, NJ: Wiley.
Google Scholar
Williamson, D. H., Lund, P., and Krebs, H. A. (1967). Redoxní stav volného nikotinamid-adenindinukleotidu v cytoplazmě a mitochondriích jater potkana. Biochem. J. 103, 514-527.
Pubmed Abstract | Pubmed Full Text | Google Scholar
.