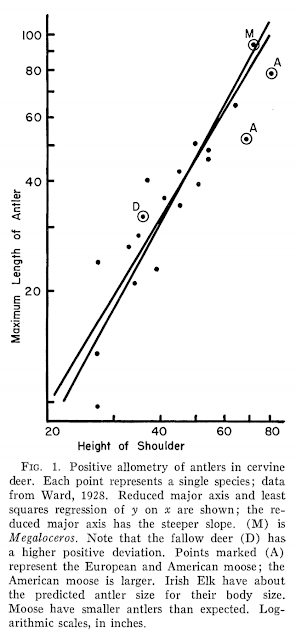
Czy poroże Megalocerosa było wyjątkowo duże? W sensie absolutnym tak, ale w sensie względnym nie: jest ono odpowiednie dla jelenia o rozmiarach jego ciała. Ten wykres z przełomowej pracy Goulda z 1974 roku jest nieco prymitywny – ani wysokość ramion, ani długość poroża nie są najlepszą miarą proporcji ciała (lepsza byłaby masa), ale to badanie dało początek bardziej świadomym i ilościowym badaniom nad ewolucją, funkcją i znaczeniem poroża olbrzymich jeleni.
Ponadto pomysły, że poroże Megalocerosa w jakiś sposób rozwijało się poza kontrolą, ignorują niuanse jego struktury i ewolucji. Nie ma wątpliwości, że – tak jak w przypadku wszystkich jeleni – samce jelenia olbrzymiego były fizjologicznie obciążone produkcją poroża. Nawet zakładając, że rozwój trwał kilka miesięcy, centymetry wzrostu były wymagane każdego dnia i wątpliwe jest, by wystarczająca ilość wapnia i fosforanu mogła być przetwarzana bezpośrednio z ich pożywienia wystarczająco szybko, by sprostać zapotrzebowaniu (Moen et al. 1999). Megaloceros zareagował standardowym przystosowaniem jeleniowatych, pożyczając materiał kostny ze swojego szkieletu, a modele fizjologiczne sugerują, że do osiągnięcia pełnego rozłożenia poroża potrzebne było do 80% dostępnego resorbowalnego wapnia – około dwa razy więcej niż u łosia (Moen et al. 1999). Zakładając dostęp do odpowiednich roślin, te resorbowane minerały mogły być zastąpione w czasie, aby odbudować silny, trwały szkielet na czas rutyny, ale osteoporoza pozostawała zagrożeniem dla jeleni, które nie były w stanie odzyskać tego materiału. Nic więc dziwnego, że poroże Megalocerosa nie jest tak masywne, jak poroże innych jeleni. Wszystkie poroża są mieszanką kości zbitej (bardzo gęstej i ciężkiej, co wiąże się z wysokim kosztem mineralnym) i kości gąbczastej (lżejszej i mniej gęstej, więc o niższym koszcie mineralnym), a Megaloceros wyewoluował z wyższego stosunku kości o niższej gęstości, by lepiej radzić sobie z fizjologicznymi wymaganiami wzrostu poroża. Jednak nawet to nie wystarczyło, by ich poroże nie stało się czasem ciężarem: w całej ich historii ewolucyjnej zaobserwowano, że średni rozmiar poroża od czasu do czasu się zmniejsza. Te reakcje są dokładnym przeciwieństwem tego, czego spodziewalibyśmy się w ortogenicznym modelu „uciekającej ewolucji”, i są dowodem na to, jak zwykłe naciski selekcyjne utrzymywały anatomię Megaloceros w zgodzie z warunkami środowiskowymi.

Poroże Megaloceros było wyjątkowym urządzeniem komunikacyjnym, wyglądającym na ogromne pod wieloma kątami, a zwłaszcza z przodu. W przeciwieństwie do innych jeleni, Megaloceros nie musiał pozować, by pokazać swoje dłonie z poroża: były one nie do przeoczenia, niezależnie od tego, jak się je widziało. Screengrab ze skanu 3D wykonanego przez National Museums Scotland, z którym można się świetnie bawić (i który stanowi użyteczne odniesienie dla artystów!).
Względnie niska wartość mineralna i rozmiar poroża Megalocerosa doprowadziły do dyskusji na temat ich funkcji: czy były one używane wyłącznie do komunikacji wizualnej? Uważa się, że obszary poroża palmate, które występują dziś u takich gatunków jak daniele i łosie, służą przede wszystkim do komunikacji, a zatem można sądzić, że dotyczyło to również Megalocerosa. Jednak podczas gdy żyjące jelenie palmate używają ruchu głowy i szyi, by zwrócić uwagę na swoje poroże (Gould 1974; Geist 1999), nakrycie głowy Megalocerosa było obracane w taki sposób, że jelenie mogły eksponować swoje ogromne dłonie po prostu stojąc w miejscu. Wykluczało to potrzebę machania 40 kg kości wokół siebie przez wiele dni, co prawdopodobnie było wspaniałą wiadomością dla tkanek szyi i głowy jeleni megalocerosów (Gould 1974). Mimo to czaszki, szyje i ramiona megalocerosów były silnie wzmocnione, by podtrzymywać poroże (dach czaszki miał 30 mm grubości! – Lister 1994) i jest prawdopodobne, że były one w stanie poruszać nim z szybkością i precyzją, być może nawet gwałtownie. Choć niektórzy autorzy (Gould 1974) uznali jelenie Megaloceros za niezdolne do siłowania się ze sobą, badania histologii ich poroża i rozkładu naprężeń sugerują, że takie pojedynki były możliwe (Lister 1994; Klinkhamer et al. 2019). Skutki zderzenia poroży muszą być jeszcze modelowane – jest to obarczone zmiennymi, które obecnie trudno oszacować – ale poroża Megaloceros dobrze radzą sobie w reżimach pchania i skręcania, pod warunkiem, że naprężenia są odczuwane głównie w ich bliższych regionach. Wydaje się, że były one szczególnie odporne na siły rotacyjne i możemy sobie wyobrazić jelenie, które w bliskiej odległości, twarzami nisko przy ziemi, ściskają swoje poroża, popychając i skręcając się nawzajem do podłogi (Lister 1994). Ich poroże jest tak długie, że siły powstające na jego końcach mogły być wzmocnione do poziomu potencjalnie łamiącego kości, więc możliwe jest, że Megaloceros unikał szczególnie gwałtownych, nieprzewidywalnych stylów walki. Ponownie, jest to zgodne z tendencjami obserwowanymi u jeleni: gdy jelenie stają się większe, a gatunki stają się bardziej towarzyskie, ich walki stają się bardziej zrytualizowane i mniej ryzykowne. Megaloceros może być ostatecznym przykładem tej korelacji (Geist 1999).
Wygląd żywota

Zdjęcie Charlesa Knighta z 1906 roku przedstawiające Megalocerosa w tradycyjnej, jeleniowatej formie. Rekonstrukcje przypominające jeleniowate są charakterystyczne dla tego rodzaju od co najmniej lat 50. XIX wieku, kiedy to Waterhouse Hawkins zrekonstruował Megaloceros dla Crystal Palace Park. Image from Wikimedia Commons, in public domain.
Although no soft-tissue Megaloceros fossils are known, a reasonable amount has been inferred about its life appearance through predictions of ecology and interpretation of Palaeolithic art (e.g. Lister 1994; Geist 1999; Guthrie 2006; also see Darren Naish’s summary at Tetrapod Zoology). Mimo, że różnią się one w niektórych szczegółach, powstał konsensus co do niektórych aspektów, w tym obecności dużego garbu na ramieniu (cecha diagnostyczna sztuki jaskiniowej Megaloceros) i przypominającej daniela wydatnej krtani. Cechy te wydają się być wspólne zarówno dla samców, jak i samic, co sugeruje ich zastosowanie poza rywalizacją samców i sygnalizacją seksualną. Kuszące jest powiązanie dużych krtani z głębokim, gardłowym nawoływaniem, podobnym do tego, jakie wydaje Dama, a skoro posiadają je przedstawiciele obu płci, to może Megaloceros był szczególnie głośnym jeleniem? Artyści paleolityczni przedstawiali garb barkowy jako duży i wybrzuszony u obu płci, więc prawdopodobnie nie reprezentuje on jedynie powiększonych mięśni barkowych i szyjnych jeleni, które przenoszą poroże. Geist (1999) spekulował, że reprezentuje on garb tłuszczowy: rezerwę energetyczną na okresy sezonowego stresu, która, będąc skoncentrowana w jednym miejscu, nie powodowałaby takiego samego ryzyka przegrzania jak rozmieszczenie tkanki tłuszczowej na całym ciele, co ma miejsce u niektórych reniferów. Bydło Zebu, nosorożce białe, żubry i bawoły są podobnie przystosowanymi gatunkami biegającymi (Geist 1999). Nie ma artystycznych dowodów na istnienie grzywy lub dzwonka, co można zaobserwować u niektórych innych dużych jeleniowatych, chociaż niektóre sztuki paleolityczne wskazują na dłuższe włosy na garbie.

Ujęcie wzorca ciała Megaloceros oparte na bardzo dosłownym odczytaniu niektórych paleolitycznych dzieł sztuki – zwróć uwagę na pręgę biegnącą od łopatki do tylnej kończyny jelenia. Garb barkowy u łani powinien być prawdopodobnie większy.
Przewidywania dotyczące ubarwienia również zostały przedstawione, choć nikt nie powinien mieć złudzeń, że mamy solidne pojęcie o kolorach i wzorach u tego gatunku. Nasze najbardziej pouczające spostrzeżenia na ten temat to sztuka jaskiniowa ukazująca ciemne kolory na garbie barkowym, które przechodzą w ciemne pasy rozciągające się na całym ciele i szyi (ukośnie na bokach od barku do okolic kolan, jak również pionowo na styku szyi i tułowia). Kolejna ciemna pręga lub pas jest odnotowana na szczycie szyi, prawdopodobnie w związku z powiększonym obszarem krtani. Inny pionowy pasek mógł obramowywać łatę na grzbiecie – cecha zgodna z Dama. Sposób interpretacji tych rysunków nie jest w 100% jasny, a rekonstrukcje całego ciała Geista (1999) i Guthriego (2006) różnią się w niektórych szczegółach. Geist uznał Megalocerosa za bladego na całym ciele z wyraźnie ciemną poprzeczną pręgą i grzbietową linią środkową, podczas gdy Guthrie przedstawił bardziej zregionalizowane ubarwienie z ciemniejszymi tylnymi częściami ciała przechodzącymi w jaśniejsze odcienie wokół łopatek, obramowane wyraźnymi pręgami. Lister (1994) był bardziej konserwatywny, odnotowując jedynie prawdopodobieństwo występowania ciemnego garbu na ramieniu.