| Lystrosaurus Fossiel bereik: Laat-Permien – Vroeg-Trias, 250-248 Ma |
|
|---|---|
 Restauratie van Lystrosaurus murrayi door kunstenaar. |
|
| Wetenschappelijke classificatie | |
|
Klasse: |
Synapsida |
|
Orde: |
Therapsida |
|
Suborde: |
Anomodontia |
|
Infra-orde: |
Dicynodontia |
|
Familie: |
Lystrosauridae |
|
Genus: |
Lystrosaurus |
|
Soorten: |
|
|
Synoniemen: |
|
Lystrosaurus (wat “schophagedis” betekent in het Grieks) is een uitgestorven geslacht van dicynodont therapsid dat leefde tijdens het Laat-Permien en het Vroeg-Trias, ongeveer 250 miljoen jaar geleden in wat nu Antarctica, India en Zuid-Afrika is. Momenteel worden 4 tot 6 soorten erkend, hoewel men in de jaren 1930 tot 1970 dacht dat het aantal soorten veel groter was. Lystrosaurus had slechts twee tanden, zoals alle andere dicynodonts, en een paar slagtandachtige hoektanden. Lystrosaurus zou een hoornige snavel hebben gehad, die werd gebruikt voor het afbijten van stukjes vegetatie. Lystrosaurus was een zwaargebouwd, plantenetend dier, ongeveer zo groot als een varken. De structuur van zijn schouders en heupgewrichten suggereren dat Lystrosaurus zich voortbewoog met een semi-spreidende gang. De voorpoten waren nog robuuster dan de achterpoten, en men denkt dat het dier een krachtige graver was die zich in holen nestelde.
Lystrosaurus was verreweg het meest voorkomende gewervelde landdier van het Vroeg-Trias, goed voor maar liefst 95% van het totale aantal individuen in sommige fossiele bedden. Vaak is gesuggereerd dat hij anatomische kenmerken had waardoor hij zich beter dan de meeste dieren kon aanpassen aan de atmosferische omstandigheden die door het uitsterven van het Perm-Trias ontstonden en die in het Vroege Trias aanhielden – lage zuurstofconcentraties en hoge kooldioxideconcentraties. Recent onderzoek suggereert echter dat deze kenmerken bij Lystrosaurus niet meer uitgesproken waren dan bij geslachten die bij de uitsterving omkwamen of geslachten die overleefden maar veel minder talrijk waren dan Lystrosaurus.
Beschrijving
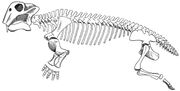
Lystrosaurus skelet diagram.
Lystrosaurus was een dicynodont therapsid ter grootte van een varken, typisch ongeveer 3 voet (0,91 m) lang en met een gewicht van ongeveer 200 pond (91 kg). In tegenstelling tot andere therapsids, hadden dicynodonts zeer korte snuiten en geen tanden behalve de slagtand-achtige bovenste hoektanden. Over het algemeen wordt aangenomen dat dicynodonten hoornachtige snavels hadden, zoals die van schildpadden, om stukken vegetatie af te scheren, die vervolgens werden vermalen op een hoornachtig secundair gehemelte wanneer de mond gesloten was. Het kaakgewricht was zwak en bewoog heen en weer met een scherende beweging, in plaats van de meer gebruikelijke zijwaartse of op en neer gaande bewegingen. Men denkt dat de kaakspieren ongewoon ver naar voren op de schedel waren bevestigd en veel ruimte op de boven- en achterkant van de schedel in beslag namen. Het gevolg was dat de ogen hoog en ver naar voren op de schedel waren geplaatst en het gezicht kort was. De onderkaak kon in opmerkelijke mate naar achteren en naar voren schuiven, waardoor het dier vegetatie in zijn bek kon brengen. Het gebruikte zijn slagtanden om wortels en knollen op te graven, die ook deel uitmaakten van zijn dieet. De neusgaten van Lystrosaurus zaten hoog op zijn diep gebogen snuit, waardoor hij zich kon voeden in het ondiepe water van zijn moeras- en moerashabitat.
Features of the skelet indicate that Lystrosaurus moved with a semi-sprewling gait. De onderste achterhoek van het scapula (schouderblad) was sterk verbeend (opgebouwd uit sterk bot), hetgeen suggereert dat beweging van het scapula bijdroeg tot de paslengte van de voorste ledematen en de zijwaartse buiging van het lichaam verminderde. De vijf heiligbeenwervels waren massief, maar niet met elkaar en met het bekken vergroeid, waardoor de rug stugger was en de zijwaartse buiging tijdens het lopen werd beperkt. Therapsiden met minder dan vijf heiligbeenwervels zouden uitgestrekte ledematen hebben gehad, zoals die van moderne hagedissen. Bij dinosauriërs en zoogdieren, die rechtopstaande ledematen hebben, zijn de heiligbeenwervels met elkaar en met het bekken vergroeid. Een steunpunt boven elk acetabulum (heupkom) zou ontwrichting van het femur (dijbeen) hebben voorkomen terwijl Lystrosaurus met een semi-spreidse gang liep. De voorpoten van Lystrosaurus waren massief, en men denkt dat Lystrosaurus een krachtige graver was.
Verspreiding en soorten
Fossielen van Lystrosaurus zijn in veel laat-Permische en vroeg-Trias aardse beenderbedden gevonden, het meest in Afrika, en in mindere mate in delen van wat nu India, China, Mongolië, Europees Rusland, en Antarctica zijn (dat toen nog niet boven de Zuidpool lag).
Soorten gevonden in Afrika
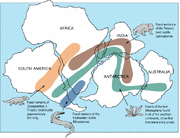
Geografische verspreiding van Lystrosaurus ( ) en hedendaagse fossielen.
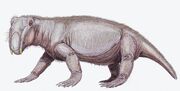
Lystrosaurus georgi
De meeste fossielen van Lystrosaurus zijn gevonden in de Balfour- en Katburg-formaties van de Karoo-regio, die grotendeels in Zuid-Afrika ligt; deze specimens bieden de beste vooruitzichten om soorten te identificeren omdat zij het talrijkst zijn en het langst bestudeerd zijn. Zoals zo vaak het geval is met fossielen, is er in de paleontologische gemeenschap discussie over het precieze aantal soorten dat in de Karoo is aangetroffen. Studies uit de jaren 1930 tot 1970 suggereerden een groot aantal (23 in één geval), maar in de jaren 1980 en 1990 werden slechts zes soorten herkend in de Karoo: L. curvatus, L. platyceps, L. oviceps, L. maccaigi, L. murrayi, en L. declivis. Een studie in 2006 bracht dat aantal terug tot vier, waarbij de fossielen die voorheen als L. platyceps en L. oviceps werden bestempeld, werden behandeld als leden van L. curvatus.
L. maccaigi is de grootste en kennelijk meest gespecialiseerde soort, terwijl L. curvatus de minst gespecialiseerde was. Een Lystrosaurus-achtig fossiel, Kwazulusaurus shakai, is ook in Zuid-Afrika gevonden. Hoewel niet tot hetzelfde geslacht behorend, lijkt K. shakai sterk op L. curvatus. Sommige paleontologen hebben daarom voorgesteld dat K. shakai mogelijk een voorouder was van of nauw verwant met de voorouders van L. curvatus, terwijl L. maccaigi uit een andere stamboom is voortgekomen.
L. maccaigi wordt alleen gevonden in sedimenten uit het Perm, en heeft blijkbaar de Perm-Trias uitstervingsgebeurtenis niet overleefd. Zijn gespecialiseerde kenmerken en plotselinge verschijning in het fossielenbestand zonder een duidelijke voorouder kunnen erop wijzen dat hij in de Karoo is geïmmigreerd uit een gebied waar geen laat-Permische sedimenten zijn gevonden.
L. curvatus wordt gevonden in een relatief smalle band van sedimenten van kort voor en na de uitsterving, en kan worden gebruikt als een benaderende marker voor de grens tussen het Perm en het Trias. Een als L. curvatus geïdentificeerde schedel is gevonden in laat-Permische sedimenten uit Zambia. Jarenlang werd gedacht dat er geen Perm-exemplaren van L. curvatus in de Karoo voorkwamen, wat tot de suggestie leidde dat L. curvatus vanuit Zambia naar de Karoo zou zijn geëmigreerd. Een nieuw onderzoek van Perm-exemplaren in de Karoo heeft er echter enkele geïdentificeerd als L. curvatus, en er is geen reden om immigratie te veronderstellen.
L. murrayi en L. declivis worden alleen in Perm sedimenten gevonden. Lystrosaurus murrayi en Lystrosaurus declivis komen alleen voor in Perm sedimenten.
Andere soorten
Fossielen van Lystrosaurus georgi zijn gevonden in de Vroegste Trias sedimenten van het Moskouse Bekken in Rusland. Hij was waarschijnlijk nauw verwant aan de Afrikaanse Lystrosaurus curvatus, die als een van de minst gespecialiseerde soorten wordt beschouwd en in zeer Laat-Permische en zeer Vroeg-Trias sedimenten is gevonden.
Geschiedenis
Dr. Elias Root Beadle, een missionaris uit Philadelphia en verwoed fossielenverzamelaar, ontdekte de eerste Lystrosaurus-schedel. Beadle schreef naar de eminente paleontoloog Othniel Charles Marsh, maar kreeg geen antwoord. Marsh’s rivaal, Edward Drinker Cope, was zeer geïnteresseerd in de vondst en beschreef en noemde Lystrosaurus in de Proceedings of the American Philosophical Society in 1870. Zijn naam is afgeleid van de Oudgriekse woorden listron “schop” en sauros “hagedis”. Marsh kocht de schedel laattijdig in mei 1871, hoewel zijn belangstelling voor een reeds beschreven specimen onduidelijk was; wellicht wilde hij Cope’s beschrijving en illustratie zorgvuldig bestuderen.
Plaattektoniek
De ontdekking van Lystrosaurus fossielen bij Coalsack Bluff in het Transantarctisch Gebergte door Edwin H. Colbert en zijn team in 1969-70 hielp de theorie van de plaattektoniek te bevestigen en de laatste twijfelaars te overtuigen, daar Lystrosaurus reeds was gevonden in het lagere Trias van zuidelijk Afrika, alsmede in India en China.
Dominantie van het Vroeg-Trias
en waarom hij de fauna van het Vroeg-Trias in zo’n ongekende mate domineerde.
Een van de meer recente theorieën is dat de Permisch-Trias uitstervingsgebeurtenis het zuurstofgehalte van de atmosfeer verminderde en het kooldioxidegehalte deed toenemen, zodat vele terrestrische soorten uitstierven omdat zij het ademen te moeilijk vonden. Men heeft daarom gesuggereerd dat Lystrosaurus overleefde en dominant werd omdat zijn ingravende levenswijze hem in staat stelde om te gaan met een atmosfeer van “muffe lucht”, en dat specifieke kenmerken van zijn anatomie deel uitmaakten van deze aanpassing: een tonvormige borstkas die plaats bood aan grote longen, korte inwendige neusgaten die een snelle ademhaling vergemakkelijkten, en hoge neurale stekels (uitsteeksels op de dorsale zijde van de wervels) die een grotere hefboomwerking gaven aan de spieren die zijn borstkas uitzetten en samentrokken. Er zijn echter zwakke punten in al deze punten: De borstkas van Lystrosaurus was in verhouding tot zijn grootte niet beduidend groter dan bij andere uitgestorven dicynodonts; hoewel Trias-dicynodonts langere neurale stekels blijken te hebben gehad dan hun Perm-tegenhangers, kan dit kenmerk eerder verband houden met houding, voortbeweging of zelfs lichaamsgrootte dan met de efficiëntie van de ademhaling; L. murrayi en L. declivis zijn veel talrijker dan andere vroeg-Trias gravers zoals Procolophon of Thrinaxodon.
De suggestie dat Lystrosaurus werd geholpen om te overleven en te domineren door semi-aquatisch te zijn heeft een vergelijkbare zwakte: hoewel amfibieën talrijker worden in de Trias sedimenten van de Karoo, zijn ze veel minder talrijk dan L. murrayi en L. declivis. De meest gespecialiseerde en de grootste dieren lopen een groter risico bij massa-extincties; dit kan verklaren waarom de niet-gespecialiseerde L. curvatus overleefde terwijl de grotere en meer gespecialiseerde L. maccaigi samen met alle andere grote Perm herbivoren en carnivoren omkwam. Hoewel Lystrosaurus over het algemeen aangepast lijkt om zich te voeden met planten vergelijkbaar met Dicroidium, die het Vroeg-Trias domineerde, kan de grotere afmeting van L. maccaigi hem gedwongen hebben om zich te voeden met de grotere leden van de Glossopteris-flora, die de uitsterving van het eind-Permien niet overleefden. Alleen de 1,5 meter lange therocephalian Moschorhinus en de grote archosauriform Proterosuchus lijken groot genoeg om op de Trias Lystrosaurus soorten te hebben gejaagd, en dit tekort aan roofdieren kan verantwoordelijk zijn geweest voor een Lystrosaurus populatie-hausse in het Vroeg-Trias. Ook is wel gesteld dat het overleven van Lystrosaurus gewoon een kwestie van geluk was.
Zie ook
- Lystrosaurus Assemblage Zone
- Lijst van synapsiden
- Evolutie van zoogdieren
- ^ “Lystrosaurus”. http://dinosaurs.about.com/od/herbivorousdinosaurs/p/lystrosaurus.htm. Op 2008-08-07 ontleend.
- ^ Cowen, R. (2000). De geschiedenis van het leven (3e ed.). Blackwell Scientific. pp. 167-68. ISBN 0-632-04444-6.
- ^ Lystrosaurus MSN Encarta encyclopedie.
- ^ a b c d e f Surkov, M.V., Kalandadze, N.N., and Benton, M.J. (June 2005). “Lystrosaurus georgi, een dicynodont uit het Neder-Trias van Rusland” (PDF). Journal of Vertebrate Paleontology 25 (2): 402-413. doi:10.1671/0272-4634(2005)0252.0.CO;2. http://palaeo.gly.bris.ac.uk/benton/reprints/2005lystrosaurus.pdf. Op 2008-08-07 ontleend.
- ^ Benton, Michael J. (2004). “Oorsprong en verwantschappen van Dinosauria”. in Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.). De Dinosauria (2e ed.). Berkeley: University of California Press. pp. 7-19. ISBN 0-520-24209-2.
- ^ a b c d e f g h i j k l m n Botha, J., and Smith, R.M.H. (2007). “Lystrosaurus species composition across the Permo-Triassic boundary in the Karoo Basin of South Africa”. Lethaia 40: 125-137. doi:10.1111/j.1502-3931.2007.00011.x. http://www3.interscience.wiley.com/journal/117996985/abstract?CRETRY=1&SRETRY=0. Afgehaald op 2008-07-02. Volledige versie online op “Lystrosaurus species composition across the Permo-Triassic boundary in the Karoo Basin of South Africa” (PDF). http://www.nasmus.co.za/PALAEO/jbotha/pdfs/Botha%20and%20Smith%202007.pdf. Op 2008-07-02 ontleend.
- ^ Grine, F.E., Forster, C.A., Cluver, M.A. & Georgi, J.A. (2006), “Cranial variability, ontogeny and taxonomy of Lystrosaurus from the Karoo Basin of South Africa”, Amniote paleobiology. Perspectives on the Evolution of Mammals, Birds, and Reptiles, University of Chicago Press, pp. 432-503
- ^ a b Wallace, David Rains (2000). De wraak van de bottenjagers: Dinosaurussen, Hebzucht en de Grootste Wetenschappelijke Vete van het Vergulde Tijdperk. Houghton Mifflin Harcourt. pp. 44-45. ISBN 0618082409.
- ^ Liddell, Henry George en Robert Scott (1980). A Greek-English Lexicon (Verkorte uitgave). Verenigd Koninkrijk: Oxford University Press. ISBN 0-19-910207-4.
- ^ Naomi Lubick, Investigating the Antarctic, Geotimes, 2005.
- ^ Erwin DH (1993). De grote Paleozoïsche crisis; Leven en dood in het Perm. Columbia University Press. ISBN 0231074670.
- ^ Cite error: Invalid
<ref>tag;er werd geen tekst verstrekt voor refs met de naamconsolation - ^ G.H Groenewald (1991). Burrow casts from the Lystrosaurus-Procolophon Assemblage-zone, Karoo Sequence, South Africa, African Protected Area Conservation and Science 34 (1): 13-22.
- ^ GM King, MA Cluver (1990). De aquatische Lystrosaurus: een alternatieve levenswijze. Historische Biologie: An International Journal of Paleobiology, 1029-2381, 4 (3):323-341.
- ^ Cite error: Ongeldige
<ref>-tag; er is geen tekst verstrekt voor refs met de naamBenton2006WhenLifeNearlyDied