| Lystrosaurus 化石の範囲。 Late Permian – Early Triassic, 250-248 Ma |
|
|---|---|
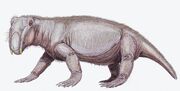
 Lystrosaurus murrayiのアーティストによる復元図です。 |
|
| 科学的分類 | |
|
Class: |
Synapsida |
|
Order.Synapsida |
|
|
Class: |
Therapsida |
|
Suborder: |
Anomodontia |
|
Inferraorder: |
Dicynodontia |
|
科: |
Lystrosauridae |
|
Genus.Of.Pirates |
Genus.Of.Pirates |
Lystrosaurus |
|
種: |
|
|
同義語: |
|
Lystrosaurus(ギリシャ語で「シャベルトカゲ」の意)は、現在の南極、インド、南アフリカで約2億5000年前の二畳紀後期から三畳紀初期に住んでいた二歯類のテラプスの絶滅属の一種です。 現在では4〜6種が確認されているが、1930年代から1970年代にかけてはもっと多いと考えられていた。 リストロサウルスは、他の二歯類と同様に2本の歯と、一対の牙のような犬歯を持っていた。 リストロサウルスは角ばったくちばしを持ち、植物のかけらを噛み切るのに使われたと考えられている。 リストロサウルスは重厚な体格の草食動物で、大きさはおよそブタである。 肩や股関節の構造から、リストロサウルスは半遊動的な歩き方をしていたようだ。 3303>
リストロサウルスは、初期三畳紀の陸生脊椎動物の中で圧倒的に多く、化石層によっては全個体の95%も占めている。 リストロサウルスは、ペルム紀から三畳紀にかけての絶滅イベントで生じた低濃度の酸素と高濃度の二酸化炭素という大気条件に、他の動物よりも適応できる解剖学的特徴を持っていたとしばしば指摘されています。 しかし、最近の研究により、リストロサウルスはこれらの特徴が、絶滅した属や、生き残ったもののリストロサウルスよりはるかに個体数の少ない属よりも顕著でなかったことが示唆されている。
説明
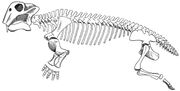
Lystrosaurus 骨格図
Lystrosaurus は豚サイズの二十歯類テラプシッドで、通常体長約 3 フィート (0.91 m) 、体重約 200ポンド (91 kg) であった。 他の獣脚類とは異なり、鼻が非常に短く、牙のような上顎を除いて歯がありませんでした。 一般に、カメのような角ばったくちばしを持ち、口を閉じたときに角ばった副口蓋で草木の切れ端をすり潰すために使っていたと考えられている。 顎の関節は弱く、一般的な横や上下の動きではなく、剪断するように前後へ動いた。 顎の筋肉が頭蓋骨の異常に前方に付着し、頭蓋骨の上部と後部に多くのスペースを取っていたと考えられている。 その結果、目は頭蓋骨の高い位置でかなり前方にあり、顔は短かった。 下あごは前後に大きく動くので、植物を口に入れることができる。 牙は根や塊茎を掘り起こすのに使われ、これも食物の一部であった。 3303>
骨格の特徴から、リストロサウルスは半匍匐前進で移動していたことがわかった。 肩甲骨の後方下角は強く骨化しており、肩甲骨の動きが前肢の歩幅に貢献し、体の横への屈曲を抑えていたと考えられる。 5つの仙椎は重厚ではあるが、骨盤と癒合していないため、背中の剛性が高く、歩行時の横屈を抑えている。 仙椎が5個より少ないセラプシス類は、現代のトカゲのようにのびのびとした手足を持っていたと考えられている。 四肢が直立する恐竜や哺乳類では、仙椎は互いに骨盤と癒合している。 リストロサウルスが半遊動歩行をしていたとき、大腿骨(太ももの骨)が脱臼しないように、それぞれの寛骨臼(股関節)の上にバットレスがあったと考えられている。 リストロサウルスの前肢は巨大で、リストロサウルスは強力な埋葬者であったと考えられている。
分布と種
リストロサウルス化石は多くのペルム紀末から三畳紀初頭の陸上骨層で発見され、アフリカで最も多く、現在のインド、中国、モンゴル、ヨーロッパロシア、南極(当時は南極を超えていなかった)の一部でわずかに見つかるだけであった。
アフリカで見つかった種
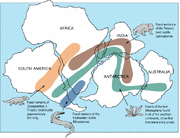
リストロサウルス ( )の地理的分布と現代の化石。
Lystrosaurus georgi
Lystrosaurus fossils have been found in Balfour and Katburg Formations of Karoo region that mostly in South Africa; these specimen has the most many and been longest time studied because the most prospect to identify species. 化石にはよくあることだが、カルー地方で発見された種が厳密に何種であるかについては、古生物学界でも議論がある。 1930年代から1970年代にかけての研究では、多くの種(一例として23種)が確認されましたが、1980年代と1990年代には、カルーではわずか6種しか確認されていません。 L. curvatus, L. platyceps, L. oviceps, L. maccaigi, L. murrayi, L. declivisである。 2006年の研究では、この数を4つに減らし、それまでL. platycepsとL. ovicepsとされていた化石をL. curvatusの仲間として扱った。
L. maccaigiは最大で明らかに最も特殊な種で、L. curvatusは最も特殊でない種であった。 また南アフリカではリストロサウルスに似た化石であるクワズサウルス・シャカイが発見されている。 K. shakaiは同属ではないが、L. curvatusと非常によく似ている。 このため、K. shakaiはL. curvatusの祖先かその近縁種であり、L. maccaigiは別の系統から発生したのではないかとする古生物学者がいる。
L.maccaigiはペルム紀の堆積物からのみ発見され、ペルム紀-三畳紀の絶滅イベントを生き延びなかったようです。 その特殊な特徴と、明らかな祖先を持たずに化石記録に突然現れたことは、ペルム紀後期の堆積物が見つかっていない地域からカルーに移住してきたことを示しているのかもしれない。
L. curvatusは絶滅の直前と直後の比較的狭い帯状の堆積物から発見され、ペルム紀と三畳紀の境界のおおよその目印とすることができる。 ザンビアのペルム紀後期堆積物からL. curvatusと同定された頭骨が発見された。 長年、カルーにはペルム紀のL. curvatusの標本はないと考えられていたため、L. curvatusはザンビアからカルーに移住したと考えられていた。 しかし、カルーにあるペルム紀の標本を再調査した結果、一部がL. curvatusと判明し、移民を仮定する必要はなくなった。
L. murrayiとL. declivisはペルム紀の堆積物からのみ見つかっている。
その他の種
ロシアのモスクワ盆地の初期三畳紀堆積物からLystrosaurus georgiの化石が発見されている。 アフリカのLystrosaurus curvatusと近縁種と思われ、最も特殊性の低い種の一つとされ、ペルム紀後期から三畳紀初期の堆積物から発見されている。
歴史
フィラデルフィアの宣教師で熱心な化石収集家だったElias Root Beadle博士が最初のリストロサウルス頭蓋骨を発見する。 ビードルは著名な古生物学者オスニエル・チャールズ・マーシュに手紙を出しましたが、返事はもらえませんでした。 マーシュのライバル、エドワード・ドリンカー・コープはこの発見に強い関心を示し、1870年の『アメリカ哲学会論文集』にリストロサウルスを記載し、命名した。 この名前は、古代ギリシャ語のリストロン「シャベル」とサウロス「トカゲ」に由来している。 マーシュは遅ればせながら1871年5月にこの頭骨を購入したが、すでに記載された標本に対する彼の関心は不明確で、コープの記述と図版を注意深く吟味したかったのかもしれない。
プレートテクトニクス
1969-70年にエドウィン・H・コルバートらのチームがトランスアンタンクティック山脈のコールサックブラフでリストロサウルス化石を発見し、プレートテクトニクス理論を確認し、最後の疑惑者を説得することができましたが、リストロサウルスはすでに南部アフリカの下部三畳紀やインド、中国で発見されていたためです。
初期の三畳紀の支配
そして、なぜこのように前例のないほど初期の三畳紀動物相を支配したのか
最近の説では、ペルム紀-三畳紀の絶滅イベントにより、大気中の酸素量が減り、二酸化炭素量が増えたために、多くの地上生物が呼吸困難になって絶滅したと言われています。 そのため、リストロサウルスは穴を掘って生活することで、”stale air “の大気に対応し、大きな肺を収容できる樽状の胸、迅速な呼吸を可能にする短い内鼻孔、胸を拡張・収縮させる筋肉に大きな力を与える高い神経突起(脊椎骨の背側にある突起)などが、その適応に貢献したと考えられてきたのである。 しかし、これらの点にはすべて弱点がある。 三畳紀の二頭類はペルム紀の二頭類よりも神経棘が長いようだが、この特徴は呼吸効率ではなく、姿勢や運動量、あるいは体格に関係している可能性があること、L. murrayiとL. また,L. murrayi と L. declivis は Procolophon や Thrinaxodon のような他の三畳紀初期埋没生物よりもはるかに多い。
リストロサウルスが半水生であることによって生存と支配に貢献したという提案も同様の弱点を持つ。カルー地方の第三紀堆積物では両生類がより豊富になるものの,それらは L. murrayi と L. declivis よりはるかに数が少なくなる。 このことは、ペルム紀の他の大型草食動物や肉食動物と一緒に、より大型で特殊化したL. maccaigiが滅び、特殊化していないL. curvatusが生き残った理由を説明することができるかもしれない。 リストロサウルスは一般に、三畳紀初期に優勢だったDicroidiumに似た植物を食べるのに適しているように見えるが、L. maccaigiのサイズが大きいため、ペルム紀末の絶滅を免れたGlossopteris植物群の大型種に頼らざるを得なかった可能性がある。 三畳紀のリストロサウルスを捕食できるのは、体長1.5メートルの獣脚類のモスコルリヌスや大型の竜脚類のプロテロスキュスだけで、この捕食者不足が三畳紀前期にリストロサウルスがブームになった理由かもしれない。 また、リストロサウルスの生存は単に運の問題であったと推測されている。
- Lystrosaurus Assemblage Zone
- List of synapsids
- 哺乳類の進化
- ^ 「リストロサウルス」(Lystrosaurus)を参照せよ。 http://dinosaurs.about.com/od/herbivorousdinosaurs/p/lystrosaurus.htm. 2008-08-07 に取得。
- ^ Cowen, R. (2000). 生命の歴史(第3版). ブラックウェルサイエンティフィック. ISBN 0-632-04444-6.
- ^ リストロサウルス MSNエンカルタ百科事典.
- ^ a b c d e f Surkov, M.V., Kalandadze, N.N., and Benton, M.J. (June 2005). “Lystrosaurus georgi, a dicynodont from the Lower Triassic of Russia” (PDF). また,”Journal of Vertebrate Paleontology” 25 (2): 402-413. 2008-08-07に取得。
- ^ Benton, Michael J. (2004). このように、「恐竜の起源と関係」、「恐竜の起源と関係」、「恐竜の起源と関係」、「恐竜の起源と関係」、「恐竜の起源と関係」、「恐竜の起源と関係」、「恐竜の起源と関係」、「恐竜の起源と関係」、「ウェシャンプール、デビッドB. The Dinosauria (2nd ed.). Berkeley: カリフォルニア大学出版局. isbn 0-520-24209-2.
- ^ a b c d e f g h i j k l m n BOTA, J., and Smith, R.M.H. (2007). 「南アフリカ共和国カルー盆地におけるペルモ三畳紀境界を越えたリストロサウルス種の構成」. http://www3.interscience.wiley.com/journal/117996985/abstract?CRETRY=1&SRETRY=0. 2008-07-02 に掲載された。 オンライン完全版 “Lystrosaurus species composition across the Permo-Triassic boundary in the Karoo Basin of South Africa” (PDF).にて公開。 http://www.nasmus.co.za/PALAEO/jbotha/pdfs/Botha%20and%20Smith%202007.pdf. 2008-07-02 に掲載。
- ^ Grine, F.E., Forster, C.A., Cluver, M.A. & Georgi, J.A. (2006), “Cranial variability, ontogeny and taxonomy of Lystrosaurus from the Karoo Basin of South Africa”, Amniote paleobiology. Perspectives on the Evolution of Mammals, Birds, and Reptiles, University of Chicago Press, pp.432-503
- ^ a b Wallace, David Rains (2000). ボーンハンターの復讐 Dinosaurs, Greed, and the Greatest Scientific Feud of the Gilded Age(恐竜、貪欲、そして金ぴか時代の最大の科学的抗争). Houghton Mifflin Harcourt. 44-45. isbn 0618082409.
- ^ Liddell, Henry George and Robert Scott (1980). A Greek-English Lexicon (Abridged Edition)(ギリシャ語-英語辞書). United Kingdom: オックスフォード大学出版局. ISBN 0-19-910207-4.
- ^ ナオミ・ルービック『南極を調べる』ジオタイムズ、2005年。
- ^ アーウィンDH (1993). 古生代の大危機;ペルム紀の生と死. コロンビア大学出版局. isbn 0231074670.
- ^ 引用エラー。 Invalid
<ref>tag;no text was provided for refs namedconsolation - ^ G.H Groenewald (1991). Lystrosaurus-Procolophon Assemblage-zone, Karoo Sequence, South Africa, African Protected Area Conservation and Science 34 (1): 13-22.
- ^ GM King, MA Cluver (1990). 水棲リストロサウルス:オルタナティブなライフスタイル. Historical Biology: An International Journal of Paleobiology, 1029-2381, 4 (3):323-341.
- ^ Cite error: 無効な
<ref>タグ。Benton2006WhenLifeNearlyDied
という名前の参照にテキストが提供されていないため。