| Lystrosaurus Fossiilivalikoima: Late Perm – Early Triassic, 250-248 Ma |
|
|---|---|
 Kuvataiteilijan restauroima Lystrosaurus murrayi. |
|
| Tieteellinen luokitus | |
|
Luokka: |
Synapsida |
|
Luokka: |
Therapsida |
|
Alaluokka: |
Anomodontia |
|
Sisäluokka: |
Dicynodontia |
|
Suku: |
Lystrosauridae |
|
Suku: |
Lystrosaurus |
|
Lajit: |
|
|
Synonyymit: |
|
Lystrosaurus (kreikaksi ”lapalisko”) on sukupuuttoon kuollut dikynodonttisten therapsidien suku, joka eli myöhäisellä permikaudella ja varhaisella triaskaudella noin 250 miljoonaa vuotta sitten nykyisen Etelämantereen, Intian ja Etelä-Afrikan alueella. Nykyisin tunnetaan 4-6 lajia, vaikka 1930-1970-luvuilla lajimäärän uskottiin olevan paljon suurempi. Lystrosauruksella oli vain kaksi hammasta, kuten kaikilla muillakin dicynodonteilla, sekä pari keihäsmäistä kulmahammasta. Lystrosauruksella uskotaan olleen kiimainen nokka, jota käytettiin kasvillisuuden palasten puremiseen. Lystrosaurus oli raskasrakenteinen, noin sian kokoinen kasvinsyöjä. Sen olkapäiden ja lonkkanivelten rakenne viittaa siihen, että Lystrosaurus liikkui puoliksi ryömivällä kävelyllä. Eturaajat olivat jopa takaraajoja järeämmät, ja eläimen uskotaan olleen voimakas kaivautuja, joka pesi pesäkoloissa.
Lystrosaurus oli varhaisen triaskauden ylivoimaisesti yleisin maaselkärankainen selkärankainen, ja sen osuus joissakin fossiilikerrostumissa oli jopa 95 prosenttia kaikista yksilöistä. Usein on esitetty, että sillä oli anatomisia ominaisuuksia, joiden ansiosta se pystyi sopeutumaan useimpia eläimiä paremmin ilmakehän olosuhteisiin, jotka syntyivät permikolmanneksen ja triaskauden välisessä sukupuuttotapahtumassa ja jotka säilyivät koko varhaisen triaskauden ajan – alhaiset happipitoisuudet ja korkeat hiilidioksidipitoisuudet. Viimeaikaiset tutkimukset viittaavat kuitenkin siihen, että nämä piirteet eivät olleet Lystrosauruksella selvempiä kuin sukupuutossa tuhoutuneilla suvuilla tai suvuilla, jotka säilyivät hengissä mutta joita oli paljon vähemmän kuin Lystrosaurusta.
Kuvaus
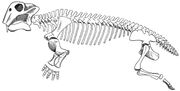
Lystrosauruksen luurankokaavio.
Lystrosaurus oli sikakokoinen dikynodonttinen therapsidi, tyypillisesti noin 3 jalan (0,91 m) pituinen ja noin 91 kg:n painoinen. Toisin kuin muilla therapsideilla, dikynodonteilla oli hyvin lyhyt kuono eikä niillä ollut hampaita lukuun ottamatta keihäsmäisiä ylähampaita. Yleisesti ajatellaan, että dicynodonteilla on ollut kilpikonnien kaltaiset sarvimaiset nokat, joiden avulla ne ovat voineet irrottaa kasvillisuuden palasia, jotka sitten jauhautuivat sarvimaisella toissijaisella suulakihalkiolla, kun suu oli suljettu. Leukanivel oli heikko, ja se liikkui eteen- ja taaksepäin leikkaavalla liikkeellä tavallisempien sivuttais- tai ylös- ja alaspäin suuntautuvien liikkeiden sijasta. On arveltu, että leukalihakset olivat kiinnittyneet kallossa epätavallisen pitkälle eteenpäin ja veivät paljon tilaa kallon ylä- ja takaosasta. Tämän seurauksena silmät asettuivat korkealle ja pitkälle kallon etuosaan, ja kasvot olivat lyhyet. Alaleuka pystyi liukumaan huomattavan paljon eteen- ja taaksepäin, minkä ansiosta eläin pystyi siirtämään kasviksia suuhunsa. Se käytti syöksyhampaitaan kaivamaan juuria ja mukuloita, jotka olivat myös osa sen ravintoa. Lystrosauruksen sieraimet olivat korkealla sen syvästi kaarevalla kuonolla, minkä ansiosta se pystyi syömään matalassa vedessä suon ja rämeen elinympäristössään.
Luuston piirteet viittaavat siihen, että Lystrosaurus liikkui puoliksi ryömivällä kävelyllä. Lapaluun (lapaluun) takimmainen alakulma oli voimakkaasti luutunut (rakentunut vahvasta luusta), mikä viittaa siihen, että lapaluun liike vaikutti osaltaan eturaajojen askelpituuteen ja vähensi vartalon sivulle taipumista. Viisi ristiluun nikamaa olivat massiivisia, mutta ne eivät olleet sulautuneet toisiinsa ja lantioon, mikä teki selästä jäykemmän ja vähensi sivuttaista taipumista eläimen kävellessä. Therapsideilla, joilla oli vähemmän kuin viisi ristinikamaa, uskotaan olleen rönsyilevät raajat, kuten nykyisillä liskoilla. Dinosauruksilla ja nisäkkäillä, joilla on pystyt raajat, ristinikamat ovat sulautuneet toisiinsa ja lantioon. Kummankin acetabulumin (lonkkamaljan) yläpuolella olevan tukipilarin uskotaan estäneen reisiluun sijoiltaanmenon, kun Lystrosaurus käveli puoliksi ryömivällä kävelyllä. Lystrosauruksen eturaajat olivat massiiviset, ja Lystrosauruksen uskotaan olleen voimakas kaivautuja.
Levinneisyys ja lajit
Lystrosauruksen fossiileja on löydetty monista myöhäispermikautisista ja varhaistriasikautisista maanpäällisistä luukerrostumista, runsaimmin Afrikasta ja vähäisemmässä määrin osista nykyistä Intiaa, Kiinaa, Mongoliaa, Eurooppalaista Venäjää ja Etelämantereelta (joka tuolloin ei ollut etelänavan yllä).
Afrikassa tavatut lajit
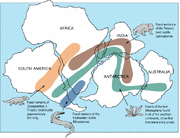
Lystrosauruksen ( ) maantieteellinen levinneisyys ja aikalaisfossiilit.
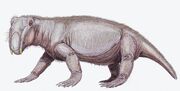
Lystrosaurus georgi
Lystrosaurus-fossiileista suurin osa on löydetty Balfourin ja Katburgin muodostumista Karoon alueelta, joka sijaitsee enimmäkseen Etelä-Afrikassa; näillä yksilöillä on parhaat mahdollisuudet lajinmääritykseen, koska niitä on eniten ja niitä on tutkittu pisimpään. Kuten niin usein fossiilien kohdalla, paleontologiyhteisössä kiistellään siitä, kuinka monta lajia Karoosta on löydetty. 1930-1970-luvuilla tehdyissä tutkimuksissa esitettiin suuri määrä (yhdessä tapauksessa 23 lajia). 1980- ja 1990-luvuilla Karoossa tunnistettiin kuitenkin vain kuusi lajia: L. curvatus, L. platyceps, L. oviceps, L. maccaigi, L. murrayi ja L. declivis. Vuonna 2006 tehdyssä tutkimuksessa määrä vähennettiin neljään, jolloin aiemmin L. platycepsiksi ja L. ovicepsiksi merkittyjä fossiileja käsiteltiin L. curvatus -lajin jäseninä.
L. maccaigi on suurin ja ilmeisesti erikoistunein laji, kun taas L. curvatus oli vähiten erikoistunut. Etelä-Afrikasta on löydetty myös Lystrosauruksen kaltainen fossiili, Kwazulusaurus shakai. Vaikka K. shakai ei kuulu samaan sukuun, se on hyvin samankaltainen kuin L. curvatus. Jotkut paleontologit ovatkin ehdottaneet, että K. shakai oli mahdollisesti L. curvatuksen esi-isien esi-isä tai läheistä sukua, kun taas L. maccaigi syntyi eri sukulinjasta.
L. maccaigi esiintyy vain permikauden sedimenteissä, eikä se ilmeisesti selvinnyt permikauden ja triaskauden välisestä sukupuuttoon kuolemisesta. Sen erikoistuneet piirteet ja yhtäkkinen esiintyminen fossiilirekisterissä ilman ilmeistä esi-isää voivat viitata siihen, että se on muuttanut Karooseen alueelta, josta ei ole löydetty myöhäispermiläisiä sedimenttejä.
L. curvatus esiintyy suhteellisen kapeassa kaistaleessa sedimenttejä hieman ennen ja jälkeen sukupuuton, ja sitä voidaan käyttää likimääräisenä merkkinä permikauden ja triaskauden välisestä rajasta. L. curvatukseksi tunnistettu kallo on löydetty Sambiasta peräisin olevista permikauden lopun sedimenteistä. Monien vuosien ajan oli luultu, että Karoossa ei ollut L. curvatus -lajin permikautisia yksilöitä, mikä johti oletuksiin, että L. curvatus olisi siirtynyt Sambiasta Karooseen. Karoon permikautisten näytteiden uudelleentarkastelu on kuitenkin osoittanut, että osa niistä on L. curvatus -lajia, eikä maahanmuuttoa ole syytä olettaa.
L. murrayi ja L. declivis esiintyvät vain permikautisissa sedimenteissä. Lystrosaurus murrayi ja Lystrosaurus declivis löytyvät vain permikautisista sedimenteistä.
Muut lajit
Lystrosaurus georgi -fossiileja on löydetty varhaisimman triaskauden sedimenteistä Moskovan altaasta Venäjältä. Se oli todennäköisesti läheistä sukua afrikkalaiselle Lystrosaurus curvatukselle, jota pidetään yhtenä vähiten erikoistuneista lajeista ja jota on löydetty hyvin myöhäisestä permikauden ja hyvin varhaisen triaskauden sedimenteistä.
Historia
Tohtori Elias Root Beadle, filadelphialainen lähetyssaarnaaja ja innokas fossiilien keräilijä, löysi ensimmäisen Lystrosaurus-kallon. Beadle kirjoitti arvostetulle paleontologille Othniel Charles Marshille, mutta ei saanut vastausta. Marshin kilpailija Edward Drinker Cope oli hyvin kiinnostunut löydöstä, ja hän kuvaili ja nimesi Lystrosauruksen vuonna 1870 ilmestyneessä American Philosophical Societyn julkaisussa Proceedings of the American Philosophical Society. Sen nimi on peräisin muinaiskreikan sanoista listron ”lapio” ja sauros ”lisko”. Marsh osti kallon myöhässä toukokuussa 1871, vaikka hänen kiinnostuksensa jo kuvattua yksilöä kohtaan oli epäselvä; hän saattoi haluta tutkia tarkkaan Copen kuvauksen ja kuvituksen.
Laattatektoniikka
Edwin H. Colbertin ja hänen tutkimusryhmänsä vuosina 1969-70 Transantarktisen vuoriston Coalsack Bluffista tekemä Lystrosaurusfossiilien löytö auttoi vahvistamaan laattatektoniikan teorian ja vakuuttamaan viimeisetkin epäilijät, sillä Lystrosaurus oli jo löydetty eteläisen Afrikan alemmasta triaskaudesta sekä Intiasta ja Kiinasta.
Varhaisen triaskauden dominointi
ja miksi se hallitsi varhaisen triaskauden eläimistöä niin ennennäkemättömän laajasti.
Yksi uudemmista teorioista on, että permitriaskauden sukupuuttotapahtuma vähensi ilmakehän happipitoisuutta ja lisäsi sen hiilidioksidipitoisuutta niin, että monet maaeläinlajit kuolivat sukupuuttoon siksi, että niiden oli liian vaikea hengittää. Onkin esitetty, että Lystrosaurus säilyi hengissä ja nousi hallitsevaksi lajiksi, koska sen kaivautuva elintapa mahdollisti sen selviytymisen ”tunkkaisesta ilmakehästä”, ja että sen anatomian erityispiirteet olivat osa tätä sopeutumista: tynnyrimainen rintakehä, johon mahtuivat suuret keuhkot, lyhyet sisäiset sieraimet, jotka helpottivat nopeaa hengittämistä, ja korkeat hermosäikeet (selkänikamien selkäpuolisten nikamien ulokkeet), jotka antoivat suuremman vipuvaikutuksen rintakehää laajeneville ja supistaville lihaksille. Kaikissa näissä kohdissa on kuitenkin heikkouksia: Lystrosauruksen rintakehä ei ollut suhteessa kokoonsa merkittävästi suurempi kuin muilla sukupuuttoon kuolleilla dicynodonteilla; vaikka triaskauden dicynodonteilla näyttäisikin olleen pidemmät hermosäikeet kuin permikauden dicynodonteilla, tämä piirre saattaa liittyä pikemminkin asentoon, liikkumiseen tai jopa ruumiin kokoon kuin hengityksen tehokkuuteen; L. murrayi ja L. declivis ovat paljon runsaampia kuin muut varhaisen triaskauden kaivautujat, kuten Procolophon tai Thrinaxodon.
Esityksessä, jonka mukaan Lystrosaurusta auttoi selviytymään ja hallitsemaan lajia se, että se oli puolivesieläin, on samanlainen heikkous: vaikka sammakkoeläimet runsastuvat Karoon triaskauden sedimenteissä, ne ovat paljon harvinaisempia kuin L. murrayi ja L. declivis. Erikoistuneimmat ja suurimmat eläimet ovat suuremmassa vaarassa joukkokuolemissa; tämä saattaa selittää, miksi erikoistumaton L. curvatus säilyi hengissä, kun taas kookkaampi ja erikoistuneempi L. maccaigi tuhoutui kaikkien muiden suurten permikauden kasvinsyöjien ja lihansyöjien mukana. Vaikka Lystrosaurus näyttää yleisesti ottaen sopeutuneen ruokailemaan samanlaisilla kasveilla kuin Dicroidium, joka hallitsi triaskauden alkupuolta, L. maccaigin suurempi koko on saattanut pakottaa sen turvautumaan Glossopteris-kasvillisuuden kookkaampiin jäseniin, jotka eivät selvinneet permikauden lopun sukupuuttoon kuolemisesta. Ainoastaan 1,5 metrin pituinen therocephalian Moschorhinus ja suuri arkkosaurusmuotoinen Proterosuchus näyttävät olleen riittävän suuria saalistamaan triaskauden Lystrosaurus-lajeja, ja tämä saalistajien puute on saattanut olla syynä Lystrosaurus-populaatioiden nousuun varhaisella triaskaudella. Lystrosauruksen selviytymisen on myös postuloitu olleen pelkkää tuuria.
Ks. myös
- Lystrosauruksen assemblaatiovyöhyke
- Luettelo synapsideista
- Nisäkkäiden evoluutio
- ^ ”Lystrosaurus”. http://dinosaurs.about.com/od/herbivorousdinosaurs/p/lystrosaurus.htm. Haettu 2008-08-07.
- ^ Cowen, R. (2000). The History of Life (Elämän historia) (3. painos). Blackwell Scientific. pp. 167-68. ISBN 0-632-04444-6.
- ^ Lystrosaurus MSN Encarta-tietosanakirja.
- ^ a b c d e f Surkov, M.V., Kalandadze, N.N., and Benton, M.J. (June 2005). ”Lystrosaurus georgi, dikynodontti Venäjän alemmasta triaskaudesta” (PDF). Journal of Vertebrate Paleontology 25 (2): 402-413. doi:10.1671/0272-4634(2005)0252.0.CO;2. http://palaeo.gly.bris.ac.uk/benton/reprints/2005lystrosaurus.pdf. Haettu 2008-08-07.
- ^ Benton, Michael J. (2004). ”Origin and relationships of Dinosauria”. in Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.). The Dinosauria (2. painos). Berkeley: University of California Press. s. 7-19. ISBN 0-520-24209-2.
- ^ a b c d e f g h i j k l m n Botha, J., and Smith, R.M.H. (2007). ”Lystrosauruksen lajikoostumus permo-triaskauden rajalla Etelä-Afrikan Karoo-altaassa”. Lethaia 40: 125-137. doi:10.1111/j.1502-3931.2007.00011.x. http://www3.interscience.wiley.com/journal/117996985/abstract?CRETRY=1&SRETRY=0. Haettu 2008-07-02. Full version online at ”Lystrosaurus species composition across the Permo-Triassic boundary in the Karoo Basin of South Africa” (PDF). http://www.nasmus.co.za/PALAEO/jbotha/pdfs/Botha%20and%20Smith%202007.pdf. Haettu 2008-07-02.
- ^ Grine, F.E., Forster, C.A., Cluver, M.A. & Georgi, J.A. (2006), ”Cranial variability, ontogeny and taxonomy of Lystrosaurus from the Karoo Basin of South Africa”, Amniote paleobiology. Perspectives on the Evolution of Mammals, Birds, and Reptiles, University of Chicago Press, pp. 432-503
- ^ a b Wallace, David Rains (2000). Luunmetsästäjien kosto: Dinosaurs, Greed, and the Greatest Scientific Feud of the Gilded Age. Houghton Mifflin Harcourt. pp. 44-45. ISBN 0618082409.
- ^ Liddell, Henry George ja Robert Scott (1980). A Greek-English Lexicon (lyhennetty painos). United Kingdom: Oxford University Press. ISBN 0-19-910207-4.
- ^ Naomi Lubick, Investigating the Antarctic, Geotimes, 2005.
- ^ Erwin DH (1993). The great Paleozoic crisis; Life and death in the Permian. Columbia University Press. ISBN 0231074670.
- ^ Cite error: Invalid
<ref>tag;no text was provided for refs namedconsolation - ^ G.H Groenewald (1991). Burrow casts from the Lystrosaurus-Procolophon Assemblage-zone, Karoo Sequence, South Africa, African Protected Area Conservation and Science 34 (1): 13-22.
- ^ GM King, MA Cluver (1990). Vedenalainen Lystrosaurus: vaihtoehtoinen elämäntapa. Historical Biology: An International Journal of Paleobiology, 1029-2381, 4 (3):323-341.
- ^ Cite error: Invalid
<ref>tag;ei annettu tekstiä viitteille nimeltäBenton2006WhenLifeNearlyDied
.