| Lystrosaurus Fossilområde: Late Permian – Early Triassic, 250-248 Ma |
|
|---|---|
 Kunstnerisk restaurering af Lystrosaurus murrayi. |
|
| Videnskabelig klassifikation | |
|
Klasse: |
Synapsida |
|
Ordning: |
Therapsida |
|
Underorden: |
Anomodontia |
|
Infraorden: |
Dicynodontia |
|
Familie: |
Lystrosauridae |
|
Genus: |
Lystrosaurus |
|
Species: |
|
|
Synonymer: |
|
Lystrosaurus (der betyder “skovlfirben” på græsk) er en uddød slægt af dicynodont therapsid, der levede i den sene Perm og tidlige Trias for omkring 250 millioner år siden i det, der nu er Antarktis, Indien og Sydafrika. På nuværende tidspunkt er der anerkendt 4 til 6 arter, selv om man fra 1930’erne til 1970’erne troede, at antallet af arter var meget højere. Lystrosaurus havde kun to tænder, som alle andre dicynodonts, samt et par stødtandslignende hjørnetænder. Lystrosaurus menes at have haft et hornagtigt næb, der blev brugt til at bide stykker af vegetation af. Lystrosaurus var et kraftigt bygget, planteædende dyr, der var omtrent på størrelse med en gris. Strukturen af dens skuldre og hofteled antyder, at Lystrosaurus bevægede sig med en halvt slyngende gangart. Forlemmerne var endnu mere robuste end baglemmerne, og dyret menes at have været en kraftig graver, der boede i huler.
Lystrosaurus var langt det mest almindelige landlevende hvirveldyr i den tidlige Trias og tegnede sig for op til 95 % af de samlede individer i nogle fossiljebunde. Det er ofte blevet foreslået, at den havde anatomiske træk, der gjorde den i stand til at tilpasse sig bedre end de fleste dyr til de atmosfæriske forhold, der blev skabt af uddøen mellem Perm og Trias, og som varede ved gennem den tidlige Trias – lave koncentrationer af ilt og høje koncentrationer af kuldioxid. Nyere forskning tyder imidlertid på, at disse egenskaber ikke var mere udtalt hos Lystrosaurus end hos slægter, der gik til grunde under uddøen, eller slægter, der overlevede, men som var langt mindre talrige end Lystrosaurus.
Beskrivelse
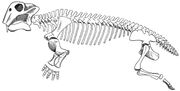
Lystrosaurus skeletdiagram.
Lystrosaurus var en dicynodont therapsid af svine-størrelse, typisk omkring 0,91 m (3 fod) lang og med en vægt på omkring 91 kg (200 pund). I modsætning til andre therapsider havde dicynodonts meget korte snuder og ingen tænder bortset fra de stødtandslignende øverste hjørnetænder. Man mener generelt, at dicynodonts har haft hornede næb som skildpadder til at skære stykker af vegetation af, som derefter blev formalet på en hornet sekundær gane, når munden var lukket. Kæbeleddet var svagt og bevægede sig frem og tilbage med en skærebevægelse i stedet for de mere almindelige sideløbende eller op- og nedadgående bevægelser. Man mener, at kæbemusklerne var anbragt usædvanligt langt fremme på kraniet og optog meget plads på kraniets over- og bagside. Som følge heraf var øjnene placeret højt og langt fremme på kraniet, og ansigtet var kort. Underkæben var i stand til at glide frem og tilbage i bemærkelsesværdig grad, hvilket gjorde det muligt for dyret at flytte vegetation ind i munden. Den brugte sine stødtænder til at grave rødder og knolde op, som også udgjorde en del af dens føde. Lystrosaurus’ næsebor var højt oppe på dens dybt buede snude, hvilket gjorde det muligt for den at æde i det lave vand i dens sump- og mosehabitat.
Kendetegn ved skelettet tyder på, at Lystrosaurus bevægede sig med en halvt slyngende gangart. Det nederste bageste hjørne af scapula (skulderbladet) var stærkt forbenet (opbygget af stærk knogle), hvilket tyder på, at scapulaens bevægelse bidrog til skridtlængden af forbenene og reducerede kroppens sidelæns bøjning i siden. De fem sakralhvirvler var massive, men ikke smeltet sammen med hinanden og med bækkenet, hvilket gjorde ryggen mere stiv og reducerede sidelæns bøjning, mens dyret gik. Therapsider med færre end fem korshvirvler menes at have haft spredte lemmer, som hos moderne øgler. Hos dinosaurer og pattedyr, som har opretstående lemmer, er korshvirvlerne smeltet sammen med hinanden og med bækkenet. Man mener, at en støtte over hvert acetabulum (hoftehule) har forhindret, at lårbenet (lårbenet) blev forskubbet, mens Lystrosaurus gik med en halvt slyngende gangart. Lystrosaurus’ forben var massive, og Lystrosaurus menes at have været en kraftig graver.
Udbredelse og arter
Lystrosaurus-fossiler er fundet i mange senpermiske og tidlige triasiske terrestriske knoglelag, mest rigeligt i Afrika og i mindre grad i dele af det, der nu er Indien, Kina, Mongoliet, det europæiske Rusland og Antarktis (som ikke var over sydpolen på det tidspunkt).
Arter fundet i Afrika
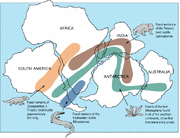
Geografisk udbredelse af Lystrosaurus ( ) og samtidige fossiler.
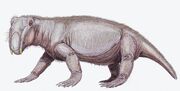
Lystrosaurus georgi
De fleste Lystrosaurus-fossiler er fundet i Balfour- og Katburg-formationerne i Karoo-regionen, som hovedsageligt ligger i Sydafrika; disse eksemplarer giver de bedste muligheder for at identificere arter, fordi de er de mest talrige og er blevet undersøgt i længst tid. Som så ofte med fossiler er der debat i det palæontologiske samfund om, hvor mange arter der præcist er fundet i Karoo. Undersøgelser fra 1930’erne til 1970’erne tydede på et stort antal (23 i et tilfælde), men i 1980’erne og 1990’erne blev der kun anerkendt seks arter i Karoo: L. curvatus, L. platyceps, L. oviceps, L. maccaigi, L. murrayi og L. declivis. En undersøgelse i 2006 reducerede dette antal til fire, idet de fossiler, der tidligere blev betegnet som L. platyceps og L. oviceps, blev behandlet som medlemmer af L. curvatus.
L. maccaigi er den største og tilsyneladende mest specialiserede art, mens L. curvatus var den mindst specialiserede. Et Lystrosaurus-lignende fossil, Kwazulusaurus shakai, er også blevet fundet i Sydafrika. Selv om K. shakai ikke er henført til den samme slægt, ligner K. shakai meget L. curvatus. Nogle palæontologer har derfor foreslået, at K. shakai muligvis var en forfader til eller nært beslægtet med forfædrene til L. curvatus, mens L. maccaigi opstod fra en anden slægt.
L. maccaigi er kun fundet i sedimenter fra Perm og har tilsyneladende ikke overlevet udryddelseshændelsen Perm-Trias. Dens specialiserede træk og pludselige optræden i fossilregistret uden en åbenlys forfader kan indikere, at den er indvandret til Karoo fra et område, hvor der ikke er fundet sedimenter fra senpermisk tid i Karoo.
L. curvatus er fundet i et relativt smalt bånd af sedimenter fra kort før og efter uddøen, og kan bruges som en omtrentlig markør for grænsen mellem Perm og Trias. Et kranie, der er identificeret som L. curvatus, er fundet i senpermiske sedimenter fra Zambia. I mange år har man troet, at der ikke fandtes permiske eksemplarer af L. curvatus i Karoo, hvilket har ført til forslag om, at L. curvatus var indvandret fra Zambia til Karoo. En ny undersøgelse af permiske eksemplarer i Karoo har imidlertid identificeret nogle af dem som L. curvatus, og der er ingen grund til at antage, at der er tale om indvandring.
L. murrayi og L. declivis er kun fundet i permiske sedimenter. Lystrosaurus murrayi og Lystrosaurus declivis findes kun i Perm-sedimenter.
Andre arter
Lystrosaurus georgi-fossiler er fundet i de tidligste triassiske sedimenter i Moskva-bækkenet i Rusland. Den var sandsynligvis nært beslægtet med den afrikanske Lystrosaurus curvatus, der betragtes som en af de mindst specialiserede arter og er fundet i sedimenter fra meget sen Perm og meget tidlig Trias.
Historie
Dr. Elias Root Beadle, en missionær fra Philadelphia og ivrig fossilsamler, opdagede det første Lystrosaurus-kranie. Beadle skrev til den eminente palæontolog Othniel Charles Marsh, men fik intet svar. Marshs rival, Edward Drinker Cope, var meget interesseret i at se fundet, og han beskrev og navngav Lystrosaurus i Proceedings of the American Philosophical Society i 1870. Dens navn er afledt af de oldgræske ord listron “skovl” og sauros “øgle”. Marsh købte kraniet sent i maj 1871, selv om hans interesse i et allerede beskrevet eksemplar var uklar; han ønskede måske at undersøge Copes beskrivelse og illustration nøje.
Pladetektonik
Den opdagelse af Lystrosaurus-fossiler ved Coalsack Bluff i de transantarktiske bjerge, som Edwin H. Colbert og hans hold gjorde i 1969-70, var med til at bekræfte teorien om pladetektonik og overbevise de sidste tvivlere, da Lystrosaurus allerede var blevet fundet i den nedre Trias i det sydlige Afrika samt i Indien og Kina.
Dominans i den tidlige Trias
og hvorfor den dominerede faunaen i den tidlige Trias i et hidtil uset omfang.
En af de nyere teorier er, at uddøen i Perm-Trias reducerede atmosfærens iltindhold og øgede dens kuldioxidindhold, så mange landlevende arter uddøde, fordi de fik for svært ved at trække vejret. Det er derfor blevet foreslået, at Lystrosaurus overlevede og blev dominerende, fordi dens gravende levevis gjorde den i stand til at klare sig i en atmosfære med “forældet luft”, og at særlige træk ved dens anatomi var en del af denne tilpasning: et tøndeformet bryst, der kunne rumme store lunger, korte indre næsebor, der gjorde det lettere at trække vejret hurtigt, og høje neuralrygge (fremspring på rygsiden af ryghvirvlerne), der gav større løftestangseffekt til de muskler, der udvidede og trak brystet sammen. Der er dog svagheder på alle disse punkter: Lystrosaurus’ brystkasse var ikke væsentligt større i forhold til dens størrelse end hos andre uddøde dicynodonts; selv om triassiske dicynodonts synes at have haft længere neuralrygge end deres permiske modstykker, kan dette træk være relateret til kropsholdning, bevægelighed eller endog kropsstørrelse snarere end til åndedrætseffektivitet; L. murrayi og L. declivis er meget mere talrige end andre tidlige Trias gravere såsom Procolophon eller Thrinaxodon.
Spørgsmålet om, at Lystrosaurus blev hjulpet til at overleve og dominere ved at være semi-akvatisk har en lignende svaghed: selv om padder bliver mere talrige i Karoo’s Trias-sedimenter, er de meget mindre talrige end L. murrayi og L. declivis. De mest specialiserede og største dyr er i større risiko ved masseudryddelser; dette kan forklare, hvorfor den ikke-specialiserede L. curvatus overlevede, mens den større og mere specialiserede L. maccaigi gik til grunde sammen med alle de andre store herbivorer og kødædere fra Perm. Selv om Lystrosaurus generelt ser ud til at være tilpasset til at æde planter svarende til Dicroidium, som dominerede den tidlige Trias, kan L. maccaigi’s større størrelse have tvunget den til at være afhængig af de større medlemmer af Glossopteris-floraen, som ikke overlevede uddøen i slutningen af Perm. Kun den 1,5 meter lange therocephalian Moschorhinus og den store archosauriform Proterosuchus synes at være store nok til at have været bytte for de triasiske Lystrosaurus-arter, og denne mangel på rovdyr kan have været skyld i et boom i Lystrosaurus-populationen i den tidlige trias. Lystrosaurus’ overlevelse er også blevet postuleret som værende simpelthen et spørgsmål om held.
Se også
- Lystrosaurus Assemblage Zone
- Liste over synapsider
- Pattedyrenes udvikling
- ^ “Lystrosaurus”. http://dinosaurs.about.com/od/herbivorousdinosaurs/p/lystrosaurus.htm. Hentet den 2008-08-07.
- ^ Cowen, R. (2000). The History of Life (3. udg.). Blackwell Scientific. pp. 167-68. ISBN 0-632-0444444-6.
- ^ Lystrosaurus MSN Encarta encyclopedia.
- ^ a b c d e f Surkov, M.V., Kalandadze, N.N., og Benton, M.J. (juni 2005). “Lystrosaurus georgi, a dicynodont from the Lower Triassic of Russia” (PDF). Journal of Vertebrate Paleontology 25 (2): 402-413. doi:10.1671/0272-4634(2005)0252.0.CO;2. http://palaeo.gly.bris.ac.uk/benton/reprints/2005lystrosaurus.pdf. Hentet den 2008-08-07.
- ^ Benton, Michael J. (2004). “Origin and relationships of Dinosauria”. in Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.). The Dinosauria (2. udg.). Berkeley: University of California Press. ISBN 0-520-24209-2.
- ^ a b c c d e f g g h i j k l l m m n Botha, J., and Smith, R.M.H. (2007). “Lystrosaurus species composition across the Permo-Triassic boundary in the Karoo Basin of South Africa”. Lethaia 40: 125-137. doi:10.1111/j.1502-3931.2007.00011.x. http://www3.interscience.wiley.com/journal/117996985/abstract?CRETRY=1&SRETRY=0. Hentet den 2008-07-02. Fuldstændig version online på “Lystrosaurus species composition across the Permo-Triassic boundary in the Karoo Basin of South Africa” (PDF). http://www.nasmus.co.za/PALAEO/jbotha/pdfs/Botha%20and%20Smith%202007.pdf. Hentet den 2008-07-02.
- ^ Grine, F.E., Forster, C.A., Cluver, M.A. & Georgi, J.A. (2006), “Cranial variability, ontogeny and taxonomy of Lystrosaurus from the Karoo Basin of South Africa”, Amniote paleobiology. Perspectives on the Evolution of Mammals, Birds, and Reptiles, University of Chicago Press, pp. 432-503
- ^ a b Wallace, David Rains (2000). The Bonehunters’ Revenge: Dinosaurs, Greed, and the Greatest Scientific Feud of the Gilded Age (Dinosaurer, grådighed og den største videnskabelige fejde i guldalderen). Houghton Mifflin Harcourt. pp. 44-45. ISBN 0618082409.
- ^ Liddell, Henry George og Robert Scott (1980). A Greek-English Lexicon (forkortet udgave). United Kingdom: Oxford University Press. ISBN 0-19-910207-4.
- ^ Naomi Lubick, Investigating the Antarctic, Geotimes, 2005.
- ^ Erwin DH (1993). The great Paleozoic crisis; Life and death in the Permian (Den store palæozoiske krise; liv og død i Perm). Columbia University Press. ISBN 0231074670.
- ^ Cite error: Invalid
<ref>tag;no text was provided for refs namedconsolation - ^ G.H Groenewald (1991). Burrow casts from the Lystrosaurus-Procolophon Assemblage-zone, Karoo Sequence, South Africa, African Protected Area Conservation and Science 34 (1): 13-22.
- ^ GM King, MA Cluver (1990). Den akvatiske Lystrosaurus: en alternativ livsstil. Historical Biology: An International Journal of Paleobiology, 1029-2381, 4 (3):323-341.
- ^ Cite error: Ugyldigt
<ref>-tag; der blev ikke angivet nogen tekst for referencer med navnetBenton2006WhenLifeNearlyDied