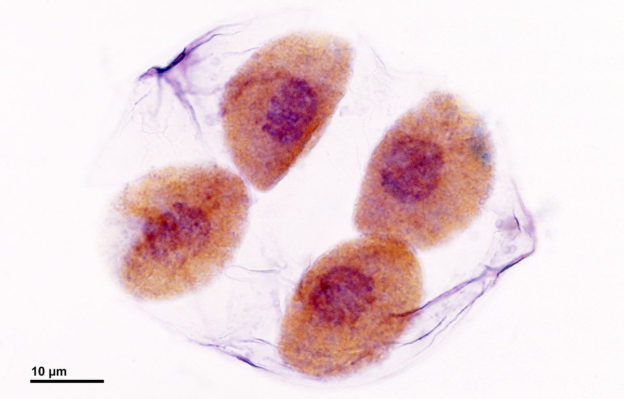
Billede: Billede: “Meiose: Deling af pollenmoderceller (ved slutningen af meiotisk deling) – Lilium-plante. Optisk mikroskopiteknik: Bright field. Forstørrelse: 3000x (for billedbredde 26 cm ~ A4-format).” af Doc. RNDr. Josef Reischig, CSc. – Forfatterens arkiv. Licens: CC BY-SA 3.0
Kromosomers grundlæggende struktur
Et kromosom er bærer af generne, de molekyler, der i sidste ende er ansvarlige for at viderebringe det arvelige sæt af oplysninger om proteinsyntese og dermed hele cellens funktion.
Et kromosom består af et par kromatider. Lignende kromosomer findes parvis og betegnes som homologe kromosomer.
En normal strukturel menneskecelle bærer to sæt, 23 par homologe kromosomer, hvilket er 46 kromosomer i alt. Dette er de diploide celler.
Den mandlige og kvindelige gametcelle, der er ansvarlig for reproduktionen, har kun ét sæt homoloide kromosomer, i alt 23, og betegnes som haploide celler.
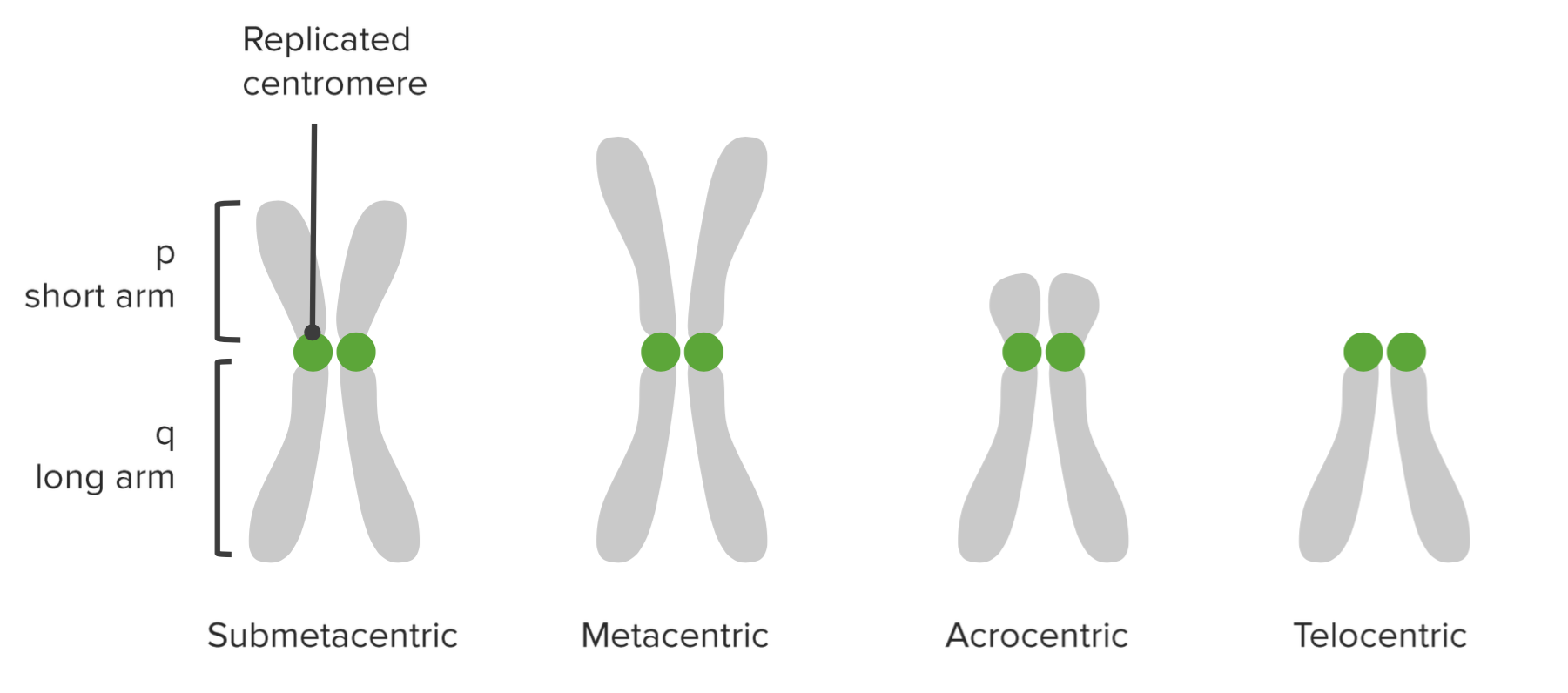
Centromeren er den del af kromosomet, hvor søsterkromatiderne er forbundet. Gennem kinetochoren fæstner spindelfibrene sig ved centromeren under celledelingen.
Centromeren afskærer hvert kromosom i to arme: den korte arm kaldes “p” og den længere “q”-arm. (p for “petit” på fransk, som betyder lille.)
Baseret på centromerets placering beskrives forskellige typer kromosomer:
| Type | Forklaring |
| Metacentrisk | Centromeret ligger i midten; p- og q-armene er lige lange. |
| Submetacentrisk | P- og q-armene er næsten lige lange. |
| Telocentrisk | Centromeren er til stede nær den ene ende; p-armen er meget lille. |
| Akrocentrisk | P-armen er lille, men lidt længere i forhold til de telocentriske kromosomer. |
Celledeling består af deling af genetisk materiale og cytogenese. Genetisk materiale kan enten gennemgå mitose eller meiose.
Mitose
I mitose giver én diploid celle anledning til to diploide celler. De to datterceller, der således opstår, er genetisk identiske med modercellen.
Tabellen viser de forskellige stadier af mitose:
| Stadie | Oplysning |
| Interfase | I celledeling veksler mitosefasen med interfasen – den tid, hvor cellen forbereder sig til delingen. Syntese af proteiner, cytoplasmaorganeller og genetisk materiale er kendetegnende for denne fase. |
| Profasen | Chromosomal kondensering og påbegyndelse af mitotisk spindeldannelse markerer profasen. |
| Metafase | Den kromosomale adskillelse efter fastgørelse af mikrotubuli ved centromeren og korrekt tilpasning langs metafasepladen eller den ækvatoriale plade sker i metafase. Metafase-kontrolpunkter sikrer en ligelig fordeling af kromosomerne ved afslutningen af den mitotiske fase af celledelingen. |
| Anafasen | Anafasen kulminerer med dannelsen af identiske datterkromosomer. Cohesiner, der binder søsterkromatiderne sammen, spaltes i denne fase. Mikrotubuli forkortes med et deraf følgende træk af et sæt nyligt dannede datterkromosomer mod modsatte ender af cellen. |
| Telofase | Afledt af det græske ord “telos”, der betyder slutningen, er telofase afslutningen på den mitotiske fase af celledelingen. Den er på mange måder en omvending af profasen. Der dannes to datterkerner med et identisk sæt kromosomer i slutningen af telofase. |
Meiose
Der er to faser i meiose, nemlig fase I og fase II.
Der sker en reduktiv deling i fase I. Chromosomal crossover, som også er et unikt træk ved fase I, fører til en udveksling af genetisk materiale mellem homologe kromosomer. Slutresultatet af meiosen er dannelsen af fire genetisk forskellige haploide celler. To haploide kønsceller fusionerer under befrugtning og genetablerer embryonets diploide natur.
Fejl i meiose som nondisjunktion er en af de mest udbredte årsager til abort og udviklingsforstyrrelser sekundært af genetisk årsag.
Fase I i meiosen kan sammenfattes således:
| Fase | Oplysning | |
| Profase I | Dette er den længste fase i meiosen. Chromosomal crossover, der fører til genetisk variation i de resulterende haploide datterceller, finder sted i profasen. Profasen er opdelt i følgende faser:
|
|
| Metafase I | Homologe kromosompar bevæger sig i denne fase langs metafasepladen. | |
| Anafase I | Homologe kromosomer bevæger sig mod modsatte poler sekundært til afkortning af kinetochore-mikrotubuli i denne fase. | |
| Telofase I | Dette stadium markerer afslutningen af den første meiotiske deling. Der dannes to datterceller, der er genetisk forskellige fra modercellen, med halvt så mange kromosomer som modercellen. Hvert kromosom består af et par kromatider. |
Fase II af meiose er identisk med mitose. Den indebærer adskillelse af søsterkromatider langs det ækvatoriale plan; ved slutningen af meiosen er der således dannet fire haploide celler.
Med denne grundlæggende indsigt i normal kromosomal anatomi og celledeling er vi klar til at gå videre til nondisjunktion.
Definition af nondisjunktion
Svigt i korrekt adskillelse af to homologe kromosomer eller søsterkromatiderne under celledeling betegnes som nondisjunktion.
Historie
Tiden vidner om opdagelsen af nondisjunktion i foråret 1910 i hænderne på Calvin Bridges og Thomas Hunt Morgan. De fandt aberrant kromosomal adfærd, mens de studerede Drosophila melanogaster kønskromosomer.
Typer af nondisjunktion
| Type | Forklaring |
| Meiotisk nondisjunktion fase I | Alle haploider afledt af den primære celle er unormale. F.eks. vil alle sædceller, der stammer fra en primær spermatocyt, have i alt 22 eller 24 kromosomer i stedet for de sædvanlige 23. |
| Meiotisk nondisjunktion fase II | Kun halvdelen af de haploider, der stammer fra primærcellen, vil være unormale. For eksempel: Hvis nondisjunktion påvirker en sekundær spermatocyt, der gennemgår meiose II, er kun halvdelen af sædcellerne unormale. |
| Mitotisk nondisjunktion | Sekundært for bruddet på spindelfibrene under metafase eller anafase resulterer mitotisk nondisjunktion i dannelsen af trisomiske og monosomiske datterceller, som giver anledning til mosaikcellelinjer hos et individ. |
Etiogenese af nondisjunktion
Molekylære mekanismer bag nondisjunktion kan kort opsummeres som følger:
| Mekanisme | Forklaring |
| Kønsspecifikke forskelle i meiose | Maternale oocytter er tilbøjelige til at have segregationsfejl, da der er dokumenteret arrest af oocytter i profase I i meiose. Som en konsekvens heraf er det et faktum, at de fleste af de menneskelige aneuploidiesyndromer er maternel afledt. |
| Aldersrelateret tab af cohesinbindinger | Cohesin er ansvarlig for fastgørelsen af spindelfibre til søsterkromatiderne og den efterfølgende normale adskillelse af samme. Længerevarende maternel oocytarrest i meiose fører til et tab af kohesinbånd; en større mulighed for ukorrekt spindelkinetochore-mikrotubulustilhæftning med deraf følgende segregationsfejl. |
| Spindle Assembly Checkpoint (SAC) funktionsfejl | SAC sikrer normal kromosomal adskillelse og tilpasning under anafasen i celledelingen. En forstyrret funktion af SAC kan føre til nondisjunktion. |
Diagnosticering af nondisjunktion
Nondisjunktion kan identificeres klinisk ved hjælp af et batteri af test som vist i nedenstående tabel:
| Test | Oplysning |
| Polar body diagnosis | Anvendes til at påvise moderligt afledte kromosomale aneuploidier. |
| Karyotypebestemmelse | En teknik, der anvender lysmikroskopi til at undersøge ufødte fosterceller, der er opnået ved fostervandsprøve. |
| Blastomerbiopsi | Indbefatter fjernelse af blastomerer fra zona pellucida for at påvise aneuploidier. Denne procedure er ikke uden risici. |
| Preimplantationsgenetisk diagnose | Anvendes hos par med en familiehistorie af genetiske lidelser, som vælger in vitro-befrugtning. |
Kliniske implikationer af nondisjunktion
Nondisjunktion resulterer i aneuploidi – en tilstand af kromosomal ubalance. Tab af et enkelt kromosom betegnes som monosomi, mens gevinst af et enkelt kromosom betegnes som trisomi. Størstedelen af de således frembragte kromosomale aberrationer er uforenelige med livet og er årsagen til størstedelen af de spontane aborter i første trimester.
Undersøgelsen af nondisjunktion afslører forbindelser mellem stigende alder hos moderen og øget chance for rekombination. Det giver også en validering af den kromosomale teori om arvelighed (Bridges 1916).
Knudsons 2-hit-hypotese for malign transformation af celler propaganderer eksistensen af en 2-trins metamorfose af den normale celle. Mens det første hit formodes at være medfødt, kan det andet hit være forårsaget af mitotisk nondisjunktion.
De få levedygtige syndromiske kromosomafvigelser kan sammenfattes således:
| Kromosomafvigelse | Forklaring |
| Monosomi | Turner syndrom (XO) er den eneste levedygtige monosomi, der er forenelig med liv hos mennesker. |
| Autosomal aneuploidi | |
| Patau syndrom (trisomi 13) | Trisomi af kromosom 13 resulterer i Patau syndrom. Det er karakteriseret ved mikrocefali, intellektuel funktionsnedsættelse, øjenproblemer, urogenitale og muskuloskeletale forstyrrelser. |
| Edwards’ syndrom (trisomi 18) | Edwards’ syndrom er kendetegnet ved tilstedeværelsen af ekstra segment-del eller hele kromosom 18. De karakteristiske træk er væksthæmning, hjertefejl, mikrognathi, alvorlig mental retardering og knyttede næver med overlappende fingre. |
| Down syndrom | Trisomi 21 er en af de mest almindelige kromosomale segregationsfejl hos mennesker. Den er notorisk kendt som “Downs syndrom” og er karakteriseret ved væksthæmning, intellektuel funktionsnedsættelse og flere neurologiske og kardiovaskulære problemer. |
| Kønskromosomaneuploidi | |
| Turners syndrom (XO) | Som allerede nævnt er dette den eneste monosomi, der er forenelig med liv hos mennesker. Det er karakteriseret ved en kort svømmehals, normal intelligens, kort statur og en højere risiko for syns- og høreproblemer. |
| Klinefelters syndrom (XXY) | Dette syndrom er karakteriseret ved tilstedeværelsen af 2 eller flere X-kromosomer hos hanner. Det er kendetegnet ved primær sterilitet, aggressiv adfærd og ofte normal intelligens med mindre tale- og læsevanskeligheder. |
| Supermales (XYY) | Karakteriseret ved XYY-genotype er denne tilstand markeret ved en forekomst på ca. 1 ud af 1.000 mandlige fødsler. Mange patienter er fænotypisk normale med større højde, lejlighedsvis aggressiv adfærd og indlæringsvanskeligheder. Det er et resultat af nondisjunktion i faderlig meiose fase II. |
| Superfemales (XXX) | Trisomi X, også betegnet som superfemales, har milde neuropsykologiske forstyrrelser. De fleste af disse er en kulmination af nondisjunktion i moderens meiose. |
| Uniparental disomi | Dette er en unik kombination af nondisjunktion, der fører til autosomal trisomi og efterfølgende tab af det uparrede kromosom, hvilket fører til eksistensen af 2 kopier af et kromosom af uniparental oprindelse. Eksempler omfatter Prader-Willi syndrom og Angelman syndrom. |
| Mosaicisme syndromer | Frølig føtal mitotisk nondisjunktion fører til den samtidige eksistens af forskellige cellelinjer hos samme individ. Hypomelanosis of Ito er et eksempel på sådanne mosaikismesyndromer. |
Summarum
Chromosomer består af par af kromatider. Homologe kromosomer findes parvis. Manglende korrekt adskillelse af homologe kromosomer eller kromatider under celledeling betegnes som nondisjunktion.
Nondisjunktion kan forekomme under meiose fase I eller fase II eller mitose.
Der findes mange test til at diagnosticere nondisjunktion.
Nondisjunktion resulterer i aneuploidi. Mens de fleste af disse kromosomale segregationsfejl fører til spontane aborter i første trimester, er få af dem forenelige med livet og fører til variable autosomale og kønskromosomale aneuploide syndromer.
Studier til medicinstudiet og bestyrelser med Lecturio.
- USMLE Step 1
- USMLE Step 2
- COMLEX Level 1
- COMLEX Level 2
- ENARM
- NEET