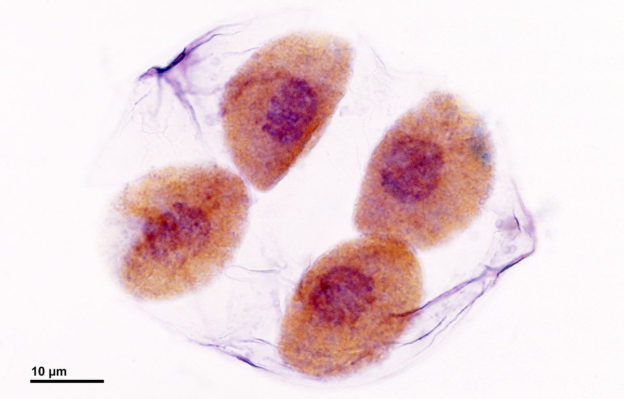
Bild: Bild: ”Meiosis: Delning av pollenmoderceller (i slutet av den meiotiska delningen) – Lilium-växt. Optisk mikroskopiteknik: Ljusfält. Förstoring: 3000x (för bildbredd 26 cm ~ A4-format).” av Doc. RNDr. Josef Reischig, CSc. – Författarens arkiv. Licens: CC BY-SA 3.0
Kromosomernas grundläggande struktur
En kromosom är bärare av gener, de molekyler som i slutändan är ansvariga för att föra vidare den ärftliga uppsättningen information om proteinsyntesen och, i sin tur, hela cellens funktion.
Varje kromosom består av ett par kromatider. Liknande kromosomer finns i par och kallas homologa kromosomer.
En normal strukturell mänsklig cell bär på två uppsättningar, 23 par homologa kromosomer, vilket är totalt 46 kromosomer. Dessa är diploida celler.
De manliga och kvinnliga könsceller som ansvarar för reproduktionen har endast en uppsättning homologa kromosomer, totalt 23 och kallas haploida celler.
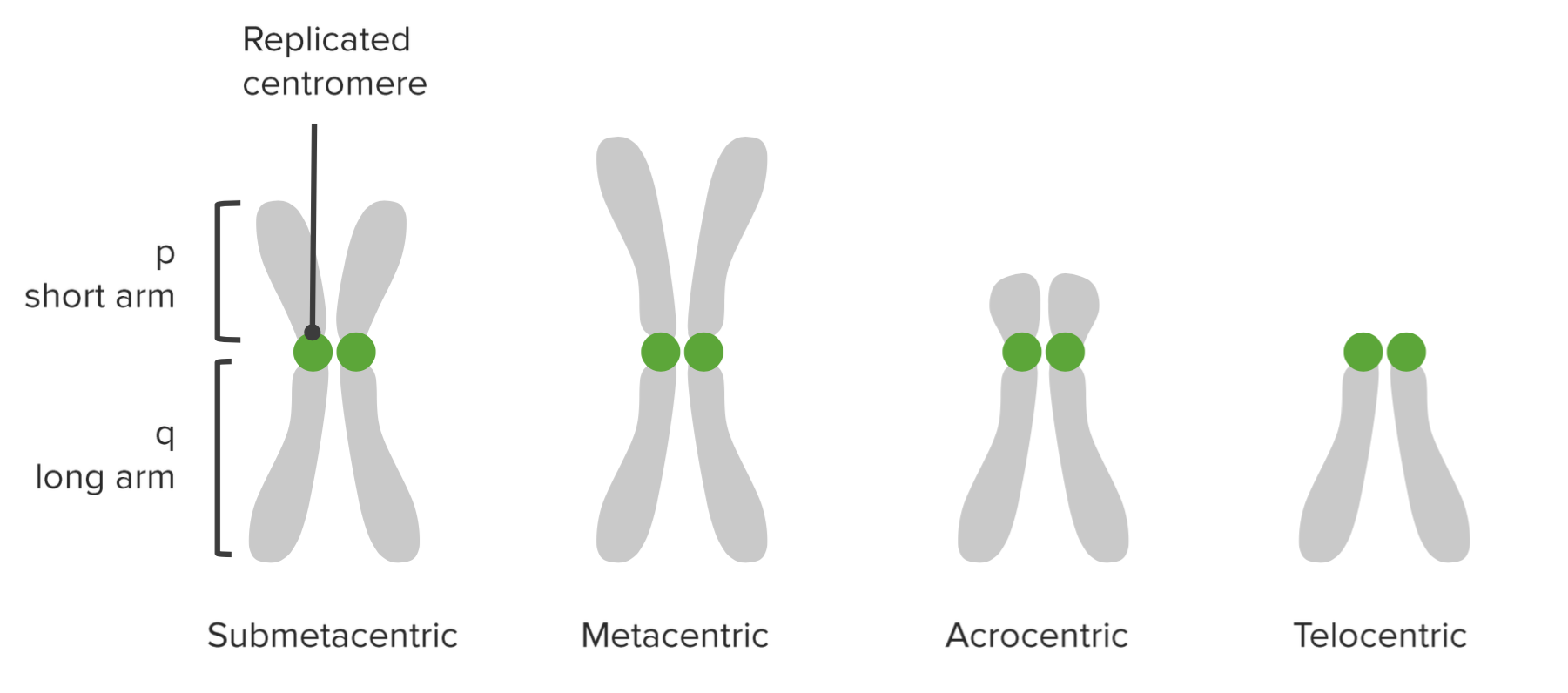
Centromeren är den del av kromosomen där systerkromatiderna är sammanlänkade. Genom kinetokorerna fäster spindelfibrerna vid centromeren under celldelningen.
Centromeren avskärmar varje kromosom i två armar: den korta armen kallas ”p” och den längre ”q”-arm. (p för ”petit” på franska som betyder liten.)
Baserat på centromerens placering beskrivs olika typer av kromosomer:
| Typ | Förklaring |
| Metacentriskt | Centromerens placering är i mitten; p- och q-arm är lika långa. |
| Submetacentric | P och q-armarna är nästan lika långa. |
| Telocentrisk | Centromer finns nära den ena änden; p-armen är mycket liten. |
| Akrocentrisk | P-armen är liten, men något längre jämfört med telocentriska kromosomer. |
Celldelning består av delning av genetiskt material och cytogenes. Genetiskt material kan antingen genomgå mitos eller meios.
Mitos
I mitos ger en diploid cell upphov till två diploida celler. De två dotterceller som på så sätt bildas är genetiskt identiska med modercellen.
Tabellen visar mitosens olika stadier:
| Stadie | Förklaring |
| Interfas | I celldelningen alternerar mitosfasen med interfasen – den tid då cellen förbereder sig för delningen. Syntesen av proteiner, cytoplasmaorganeller och genetiskt material är utmärkande för denna fas. |
| Profas | Kromosomkondensering och initiering av mitotisk spindelbildning markerar profasen. |
| Metafas | Kromosomal separation efter fastsättning av mikrotubuli vid centromeren och korrekt inriktning längs metafasplattan eller ekvatorialplattan sker i metafas. Metafasens kontrollpunkter säkerställer en jämn fördelning av kromosomerna i slutet av celldelningens mitotiska fas. |
| Anafas | Anafasen kulminerar i bildandet av identiska dotterkromosomer. Kohesiner som binder systerkromatiderna samman klyvs i denna fas. Mikrotubulerna förkortas med en resulterande dragning av en uppsättning nybildade dotterkromosomer mot motsatta ändar av cellen. |
| Telofas | Avledd från det grekiska ordet ”telos”, som betyder slutet, är telofasen slutet på den mitotiska fasen av celldelningen. Den är på många sätt en omvändelse av profasen. Två dotterkärnor med en identisk uppsättning kromosomer bildas i slutet av telofasen. |
Meios
Det finns två faser i meiosen, nämligen fas I och fas II.
Den reduktiva delningen sker i fas I. Kromosomal korsning, som också är ett unikt kännetecken för fas I, leder till ett utbyte av genetiskt material mellan homologa kromosomer. Slutresultatet av meiosen är bildandet av fyra genetiskt skilda haploida celler. Två haploida könsceller som fusioneras under befruktningen återställer embryots diploida natur.
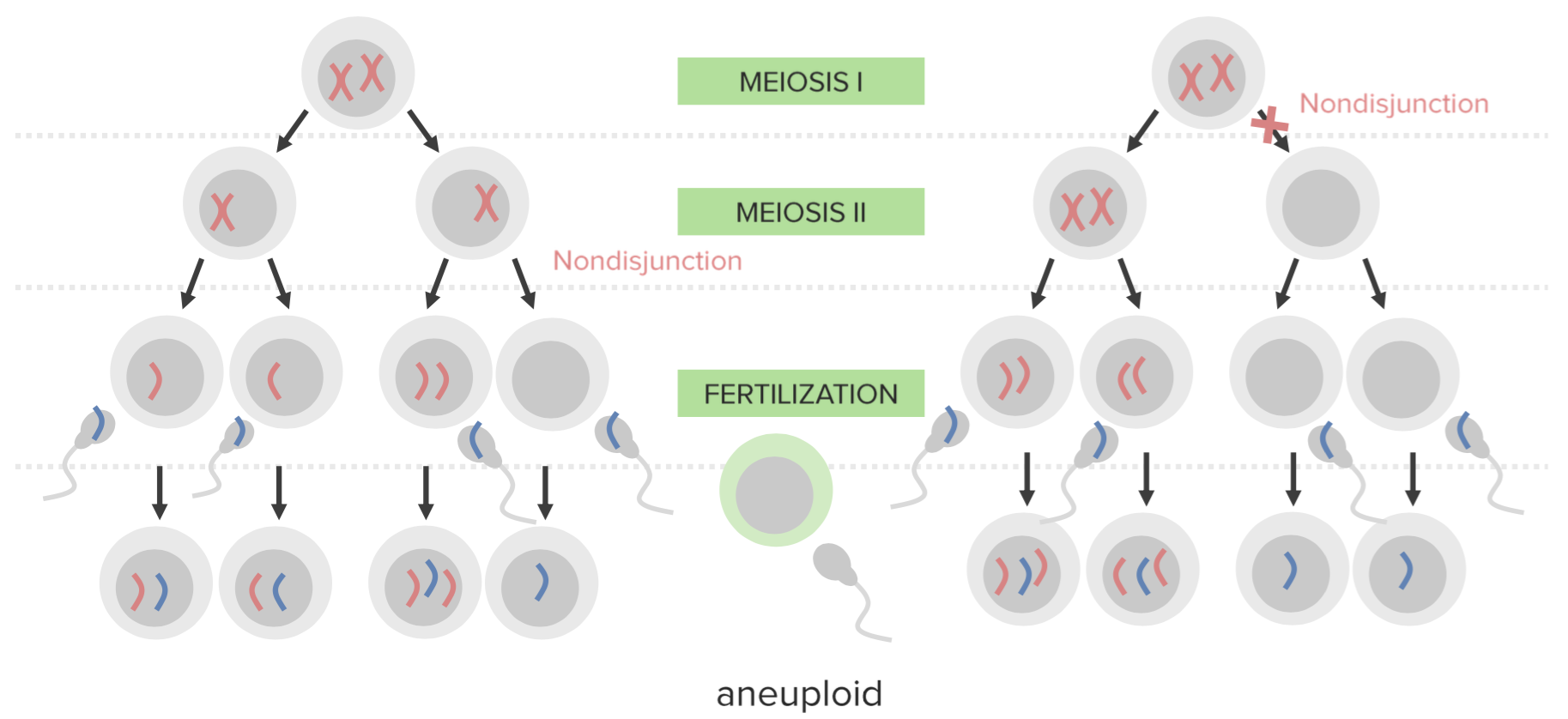
Fel i meiosen som nondisjunktion är en av de vanligaste orsakerna till missfall och utvecklingsstörningar sekundärt till en genetisk orsak.
Fas I i meiosen kan sammanfattas på följande sätt:
| Fas | Förklaring |
| Profas I | Detta är den längsta fasen i meiosen. Kromosomala korsningar som leder till genetisk variation i de resulterande haploida dottercellerna äger rum i profasen. Profasen är indelad i följande stadier:
|
| Metafas I | Homologa kromosompar rör sig längs metafasplattan i detta stadium. |
| Anafas I | Homologiska kromosomer rör sig mot motsatta poler sekundärt till förkortningen av kinetokore-mikrotubuli i denna fas. |
| Telofas I | Detta skede markerar slutet på den första meiotiska divisionen. Två dotterceller som är genetiskt skilda från modercellen bildas med halva antalet kromosomer. Varje kromosom består av ett par kromatider. |
Fas II i meiosen är identisk med mitos. Det innebär att systerkromatiderna separeras längs ekvatorialplanet; i slutet av meiosen bildas således fyra haploida celler.
Med denna grundläggande insikt om normal kromosomal anatomi och celldelning är vi redo att gå vidare till nondisjunktion.
Definition av nondisjunktion
Svikande av korrekt separation av två homologa kromosomer eller systerkromatiderna under celldelningen benämns nondisjunktion.
Historia
Tiden vittnar om att nondisjunktionen upptäcktes våren 1910 av Calvin Bridges och Thomas Hunt Morgan. De fann avvikande kromosomalt beteende när de studerade Drosophila melanogasters könskromosomer.
Typer av nondisjunktion
| Typ | Förklaring |
| Meiotisk nondisjunktion fas I | Alla haploider som härstammar från primärcellen är onormala. Till exempel kommer alla spermier som härstammar från en primär spermatocyt att ha totalt 22 eller 24 kromosomer i stället för de vanliga 23. |
| Meiotisk nondisjunktion fas II | Bara hälften av de haploider som härstammar från primärcellen kommer att vara onormala. Till exempel: Om nondisjunktion påverkar en sekundär spermatocyt som genomgår meiosis II är endast hälften av spermierna onormala. |
| Mitotisk nondisjunktion | Sekundärt till att spindelfibrerna bryts under metafas eller anafas resulterar mitotisk nondisjunktion i bildandet av trisomiska och monosomiska dotterceller, som ger upphov till mosaikcellslinjer hos en individ. |
Etiogenes av nondisjunktion
Molekylära mekanismer bakom nondisjunktion kan kortfattat sammanfattas enligt följande:
| Mekanism | Förklaring |
| Könsspecifika skillnader i meiosen | Maternala oocyter är benägna att ha segregeringsfel eftersom det finns dokumenterat att oocyter har ett stopp i profas I av meiosen. Som en följd av detta kvarstår det faktum att de flesta av de mänskliga aneuploidiesyndromen är maternellt härledda. |
| Åldersrelaterad förlust av kohesinbindningar | Kohesin är ansvarigt för att spindelfibrerna fäster vid systerkromatiderna och den efterföljande normala separationen av dessa. Långvarigt moderligt oocytstopp i meios leder till en förlust av kohesinbindningar; större möjlighet till felaktig spindelkinetokor-mikrotubuliinfästning med resulterande segregationsfel. |
| Spindle Assembly Checkpoint (SAC) felfunktion | SAC säkerställer normal kromosomal separation och anpassning under anafasen i celldelningen. Felaktig funktion av SAC kan leda till nondisjunktion. |
Diagnos av nondisjunktion
Nondisjunktion kan identifieras kliniskt med hjälp av ett batteri av tester enligt tabellen nedan:
| Test | Förklaring |
| Polkroppsdiagnos | Används för att upptäcka kromosomala aneuploidier som härrör från modern. |
| Karyotypering | En teknik som använder ljusmikroskopi för att studera ofödda fosterceller som erhållits genom fosterprovtagning. |
| Blastomerbiopsi | Involverar avlägsnande av blastomerer från zona pellucida för att påvisa aneuploidier. Detta förfarande är inte utan risker. |
| Preimplantatorisk genetisk diagnos | Används hos par med en familjehistoria av genetiska sjukdomar som väljer in-vitrofertilisering. |
Kliniska implikationer av nondisjunktion
Nondisjunktion resulterar i aneuploidi – ett tillstånd av kromosomala obalanser. Förlust av en enda kromosom kallas monosomi, medan vinst av en enda kromosom kallas trisomi. Majoriteten av de sålunda producerade kromosomavvikelserna är oförenliga med livet och är orsaken till majoriteten av spontana aborter i första trimestern.
Studien av nondisjunktion avslöjar kopplingar mellan stigande ålder hos modern och ökade chanser till rekombination. Det ger också en validering av den kromosomala teorin om ärftlighet (Bridges 1916).
Knudsons 2 hit-hypotes för malign omvandling av celler propagerar existensen av en metamorfos i två steg av den normala cellen. Medan den första träffen antas vara medfödd kan den andra träffen orsakas av mitotisk nondisjunktion.
De få livskraftiga syndromiska kromosomavvikelserna kan sammanfattas på följande sätt:
| Kromosomavvikelse | Förklaring |
| Monosomi | Turners syndrom (XO) är den enda livskraftiga monosomin som är förenlig med liv hos människor. |
| Autosomal aneuploidi | |
| Patau syndrom (trisomi 13) | Trisomi av kromosom 13 leder till Patau syndrom. Det kännetecknas av mikrocefali, intellektuell funktionsnedsättning, okulära problem, urogenitala och muskuloskeletala störningar. |
| Edwards syndrom (trisomi 18) | Edwards syndrom kännetecknas av förekomst av extra segment-del eller hela kromosom 18. Dess karakteristiska kännetecken är tillväxthämning, hjärtfel, mikrognathi, allvarlig mental retardation och knutna nävar med överlappande fingrar. |
| Downs syndrom | Trisomi 21 är ett av de vanligaste kromosomala segregationsfelen hos människor. Det är notoriskt känt som ”Downs syndrom” och kännetecknas av tillväxthämning, intellektuell funktionsnedsättning och flera neurologiska och kardiovaskulära problem. |
| Könskromosomaneuploidi | |
| Turners syndrom (XO) | Som redan nämnts är detta den enda monosomin som är förenlig med liv hos människor. Det kännetecknas av en kort svävad nacke, normal intelligens, kortväxthet och högre risk för syn- och hörselproblem. |
| Klinefelters syndrom (XXY) | Detta syndrom kännetecknas av förekomsten av 2 eller fler X-kromosomer hos män. Det kännetecknas av primär sterilitet, aggressivt beteende och ofta normal intelligens med mindre tal- och lässvårigheter. |
| Supermales (XYY) | Karaktäriseras av XYY-genotypen, detta tillstånd kännetecknas vid en incidens på cirka 1 av 1 000 manliga födslar. Många patienter är fenotypiskt normala med större längd, ibland aggressivt beteende och inlärningssvårigheter. Det är ett resultat av nondisjunktion i faderns meiosfas II. |
| Superfemales (XXX) | Trisomi X, även kallad superfemales, har milda neuropsykologiska störningar. Majoriteten av dessa är en kulmination av nondisjunktion i moderns meios. |
| Uniparental disomi | Detta är en unik kombination av nondisjunktion som leder till autosomal trisomi och efterföljande förlust av den oparade kromosomen, vilket leder till förekomsten av 2 kopior av en kromosom av uniparentalt ursprung. Exempel är Prader-Willis syndrom och Angelmans syndrom. |
| Mosaikism-syndrom | Från tidigt foster leder mitotisk nondisjunktion till samtidig existens av olika cellinjer hos samma individ. Hypomelanosis of Ito är en illustration av sådana mosaikismsyndrom. |
Sammanfattning
Kromosomer består av par av kromatider. Homologa kromosomer finns i par. Om homologa kromosomer eller kromatider inte separeras korrekt under celldelningen kallas detta för nondisjunktion.
Nondisjunktion kan inträffa under meiosens fas I eller fas II, eller mitos.
Det finns många tester tillgängliga för att diagnostisera nondisjunktion.
Nondisjunktion resulterar i aneuploidi. Medan de flesta av dessa kromosomala segregationsfel leder till spontana aborter i första trimestern, är några få förenliga med livet och leder till varierande autosomala och könskromosomala aneuploida syndrom.
Studiera för läkarutbildningar och nämnder med Lecturio.
- USMLE steg 1
- USMLE steg 2
- COMLEX nivå 1
- COMLEX nivå 2
- ENARM
- NEET