| Lystrosaurus Fossilområde: Sen permisk tid – tidig trias, 250-248 Ma |
|
|---|---|
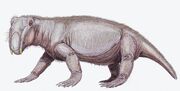
 Konstnärsrestaurering av Lystrosaurus murrayi. |
|
| Vetenskaplig klassificering | |
|
Klass: |
Synapsida |
|
Ordning: |
Therapsida |
|
Underordning: |
Anomodontia |
|
Infraordning: |
Dicynodontia |
|
Familj: |
Lystrosauridae |
|
Genus: |
Lystrosaurus |
|
Arter: |
|
|
Synonymer: |
|
Lystrosaurus (som betyder ”spadeödla” på grekiska) är ett utdött släkte av dicynodont therapsid som levde under senpermian och tidig trias, för cirka 250 miljoner år sedan, i det som nu är Antarktis, Indien och Sydafrika. För närvarande erkänns 4 till 6 arter, även om man från 1930-talet till 1970-talet trodde att antalet arter var mycket högre. Lystrosaurus hade endast två tänder, som alla andra dicynodonts, samt ett par tuskliknande hörntänder. Lystrosaurus tros ha haft en hornig näbb som användes för att bita av växtdelar. Lystrosaurus var ett kraftigt byggt, växtätande djur, ungefär lika stort som en gris. Strukturen på dess axlar och höftleder tyder på att Lystrosaurus rörde sig med en halvt spretig gång. Frambenen var ännu mer robusta än bakbenen, och djuret tros ha varit en kraftfull grävare som häckade i hålor.
Lystrosaurus var det överlägset vanligaste landlevande ryggradsdjuret i tidig trias, och stod för så mycket som 95 % av de totala individerna i vissa fossilbäddar. Det har ofta föreslagits att den hade anatomiska egenskaper som gjorde att den bättre än de flesta djur kunde anpassa sig till de atmosfäriska förhållanden som skapades av utdöendet mellan perm och trias och som kvarstod under tidig trias – låga koncentrationer av syre och höga koncentrationer av koldioxid. Ny forskning tyder dock på att dessa egenskaper inte var mer uttalade hos Lystrosaurus än hos släkten som försvann vid utdöendet eller släkten som överlevde men var mycket mindre vanliga än Lystrosaurus.
Beskrivning
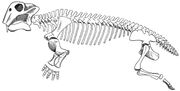
Lystrosaurus skelettdiagram
Lystrosaurus var en grisstor dicynodont therapsid, vanligen cirka 0,91 m lång och vägde cirka 91 kg. Till skillnad från andra therapsider hade dicynodonts mycket korta nosar och inga tänder förutom de tuskliknande övre hörntänderna. Dicynodonts anses i allmänhet ha haft horniga näbbar som sköldpaddor, för att klippa av vegetationsbitar som sedan maldes på en hornig sekundär gom när munnen var stängd. Käkleden var svag och rörde sig bakåt och framåt med en klippande rörelse, i stället för de vanligare rörelserna i sidled eller uppåt och nedåt. Man tror att käkmusklerna var fäst ovanligt långt fram på skallen och tog upp mycket utrymme på skallens ovansida och baksida. Detta resulterade i att ögonen satt högt och långt fram på skallen och att ansiktet var kort. Underkäken kunde glida fram och tillbaka i en anmärkningsvärd utsträckning, vilket gjorde det möjligt för djuret att föra in vegetation i munnen. Den använde sina betar för att gräva upp rötter och knölar, som också utgjorde en del av dess föda. Lystrosaurus näsborrar låg högt upp på den djupt böjda nosen, vilket gjorde det möjligt för den att livnära sig i det grunda vattnet i dess träsk- och sumpmiljö.
Skelettets egenskaper tyder på att Lystrosaurus rörde sig med en halvt sprungande gång. Det nedre bakre hörnet av scapula (skulderbladet) var starkt förbenat (byggt av starkt ben), vilket tyder på att rörelsen av scapula bidrog till frambenens steglängd och minskade kroppens böjning i sidled. De fem sakralkotorna var massiva men inte sammansvetsade med varandra och med bäckenet, vilket gjorde ryggen mer stel och minskade sidoböjningen när djuret gick. Therapsider med färre än fem sakrala ryggkotor tros ha haft utspridda lemmar, som hos moderna ödlor. Hos dinosaurier och däggdjur, som har upprättstående lemmar, är sakralkotorna sammanfogade med varandra och med bäckenet. Man tror att en stöttning ovanför varje acetabulum (höftskål) har förhindrat att lårbenet (lårbenet) rubbades när Lystrosaurus gick med en halvt sprungande gång. Lystrosaurus framben var massiva, och Lystrosaurus tros ha varit en kraftfull grävare.
Utbredning och arter
Fossil av Lystrosaurus har hittats i många sena permiska och tidiga triasiska terrestriska benlager, mest rikligt i Afrika och i mindre utsträckning i delar av vad som nu är Indien, Kina, Mongoliet, det europeiska Ryssland och Antarktis (som vid den tiden inte låg över Sydpolen).
Arter som hittats i Afrika
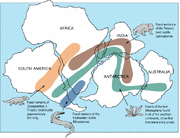
Geografisk utbredning av Lystrosaurus ( ) och samtida fossiler.
Lystrosaurus georgi
De flesta Lystrosaurus-fossilerna har hittats i Balfour- och Katburg-formationerna i Karoo-regionen, som mestadels ligger i Sydafrika; dessa exemplar erbjuder de bästa utsikterna för att identifiera arter, eftersom de är de talrikaste och har studerats under längst tid. Som så ofta när det gäller fossil finns det debatt inom paleontologkåren om exakt hur många arter som har hittats i Karoo. Studier från 1930- till 1970-talen tyder på ett stort antal (23 i ett fall), men på 1980- och 1990-talen kände man bara igen sex arter i Karoo: L. curvatus, L. platyceps, L. oviceps, L. maccaigi, L. murrayi och L. declivis. En studie från 2006 reducerade detta antal till fyra, och behandlade de fossil som tidigare betecknats som L. platyceps och L. oviceps som medlemmar av L. curvatus.
L. maccaigi är den största och uppenbarligen mest specialiserade arten, medan L. curvatus var den minst specialiserade. Ett Lystrosaurus-liknande fossil, Kwazulusaurus shakai, har också hittats i Sydafrika. Även om K. shakai inte hänförs till samma släkte är den mycket lik L. curvatus. Vissa paleontologer har därför föreslagit att K. shakai möjligen var en förfader till eller nära besläktad med L. curvatus förfäder, medan L. maccaigi uppstod från en annan släktlinje.
L. maccaigi finns endast i sediment från permperioden och överlevde uppenbarligen inte utdöendet mellan perm och trias. Dess specialiserade egenskaper och plötsliga uppkomst i fossilregistret utan en uppenbar förfader kan tyda på att den invandrat till Karoo från ett område där senpermianska sediment inte har hittats.
L. curvatus finns i ett relativt smalt band av sediment från strax före och efter utdöendet och kan användas som en ungefärlig markör för gränsen mellan permisk och triassisk period. Ett kranium som identifierats som L. curvatus har hittats i sent permiska sediment från Zambia. Under många år har man trott att det inte fanns några permiska exemplar av L. curvatus i Karoo, vilket ledde till förslag om att L. curvatus invandrat från Zambia till Karoo. En ny undersökning av permiska exemplar i Karoo har dock visat att några av dem är L. curvatus, och det finns ingen anledning att anta att det rör sig om invandring.
L. murrayi och L. declivis finns endast i permiska sediment. Lystrosaurus murrayi och Lystrosaurus declivis finns endast i permiska sediment.
Andra arter
Lystrosaurus georgi-fossil har hittats i de tidigaste triassiska sedimenten i Moskvabassängen i Ryssland. Den var troligen nära besläktad med den afrikanska Lystrosaurus curvatus, som anses vara en av de minst specialiserade arterna och har hittats i mycket sen permiska och mycket tidiga triassiska sediment.
Historia
Dr. Elias Root Beadle, en missionär från Philadelphia och ivrig fossilsamlare, upptäckte det första Lystrosaurus-kraniet. Beadle skrev till den framstående paleontologen Othniel Charles Marsh, men fick inget svar. Marshs rival, Edward Drinker Cope, var mycket intresserad av att se fyndet och beskrev och namngav Lystrosaurus i Proceedings of the American Philosophical Society 1870. Namnet kommer från de forntida grekiska orden listron ”spade” och sauros ”ödla”. Marsh köpte kraniet sent i maj 1871, även om hans intresse för ett redan beskrivet exemplar var oklart; han kanske ville granska Copes beskrivning och illustration noggrant.
Plattentektonik
Fyndet av Lystrosaurus-fossil vid Coalsack Bluff i de transantarktiska bergen av Edwin H. Colbert och hans team 1969-70 bidrog till att bekräfta teorin om plattentektonik och övertyga de sista tvivlarna, eftersom Lystrosaurus redan hade hittats i nedre trias i södra Afrika samt i Indien och Kina.
Dominans i tidig trias
och varför den dominerade faunan i tidig trias i en sådan oöverträffad omfattning.
En av de nyare teorierna är att utdöendehändelsen permisk-trias minskade atmosfärens syrehalt och ökade dess koldioxidhalt, vilket ledde till att många landlevande arter dog ut för att de hade för svårt att andas. Det har därför föreslagits att Lystrosaurus överlevde och blev dominerande eftersom dess grävande levnadssätt gjorde att den kunde klara av en atmosfär med ”förbrukad luft”, och att särskilda drag i dess anatomi var en del av denna anpassning: en tunn bröstkorg som rymde stora lungor, korta inre näsborrar som underlättade snabb andning, och höga neuralryggar (utskjutande delar på ryggkotornas ryggsida) som gav större hävstångseffekt åt musklerna som expanderade och drog ihop bröstkorgens bröstkorg. Det finns dock svagheter i alla dessa punkter: Även om de triassiska dikynodonterna tycks ha haft längre nervstjärnor än sina permiska motsvarigheter, kan denna egenskap vara relaterad till hållning, förflyttning eller till och med kroppsstorlek snarare än till andningseffektivitet. declivis är mycket rikligare än andra grävmaskiner från tidig trias, t.ex. Procolophon eller Thrinaxodon.
Suget att Lystrosaurus fick hjälp att överleva och dominera genom att vara halvt vattenlevande har en liknande svaghet: även om amfibier blir rikligare i Karoo’s triassiska sediment, så är de mycket mindre talrika än L. murrayi och L. declivis. De mest specialiserade och största djuren löper större risk vid massutrotningar; detta kan förklara varför den icke-specialiserade L. curvatus överlevde medan den större och mer specialiserade L. maccaigi gick under tillsammans med alla andra stora permiska växtätare och köttätare. Även om Lystrosaurus i allmänhet ser ut att vara anpassad för att livnära sig på växter som liknar Dicroidium, som dominerade den tidiga trias, kan L. maccaigis större storlek ha tvingat den att förlita sig på de större medlemmarna av Glossopteris-floran, som inte överlevde utdöendet i slutet av permiskan. Endast den 1,5 meter långa therocephalian Moschorhinus och den stora archosauriformen Proterosuchus tycks vara tillräckligt stora för att ha gjort byten på de triasiska Lystrosaurus-arterna, och denna brist på rovdjur kan ha varit orsaken till att Lystrosaurus-populationen ökade kraftigt under den tidiga trias. Lystrosaurus överlevnad har också postulerats som att den helt enkelt var en fråga om tur.
Se även
- Lystrosaurus Assemblage Zone
- Lista över synapsider
- Däggdjurs utveckling
- ^ ”Lystrosaurus”. http://dinosaurs.about.com/od/herbivorousdinosaurs/p/lystrosaurus.htm. Hämtad 2008-08-07.
- ^ Cowen, R. (2000). The History of Life (3:e upplagan). Blackwell Scientific. ISBN 0-632-0444444-6.
- ^ Lystrosaurus MSN Encarta encyclopedia.
- ^ a b c d e f Surkov, M.V., Kalandadze, N.N. och Benton, M.J. (juni 2005). ”Lystrosaurus georgi, a dicynodont from the Lower Triassic of Russia” (PDF). Journal of Vertebrate Paleontology 25 (2): 402-413. doi:10.1671/0272-4634(2005)0252.0.CO;2. http://palaeo.gly.bris.ac.uk/benton/reprints/2005lystrosaurus.pdf. Hämtad 2008-08-07.
- ^ Benton, Michael J. (2004). ”Origin and relationships of Dinosauria”. i Weishampel, David B.; Dodson, Peter; och Osmólska, Halszka (red.). The Dinosauria (2nd ed.). Berkeley: University of California Press. ISBN 0-520-24209-2.
- ^ a b c d e f g h i j k l m n Botha, J., and Smith, R.M.H. (2007). ”Lystrosaurus species composition across the Permo-Triassic boundary in the Karoo Basin of South Africa”. Lethaia 40: 125-137. doi:10.1111/j.1502-3931.2007.00011.x. http://www3.interscience.wiley.com/journal/117996985/abstract?CRETRY=1&SRETRY=0. Hämtad 2008-07-02. Full version online på ”Lystrosaurus species composition across the Permo-Triassic boundary in the Karoo Basin of South Africa” (PDF). http://www.nasmus.co.za/PALAEO/jbotha/pdfs/Botha%20and%20Smith%202007.pdf. Hämtad 2008-07-02.
- ^ Grine, F.E., Forster, C.A., Cluver, M.A. & Georgi, J.A. (2006), ”Cranial variability, ontogeny and taxonomy of Lystrosaurus from the Karoo Basin of South Africa”, Amniote paleobiology. Perspectives on the Evolution of Mammals, Birds, and Reptiles, University of Chicago Press, pp. 432-503
- ^ a b Wallace, David Rains (2000). The Bonehunters’ Revenge: Dinosaurs, Greed, and the Greatest Scientific Feud of the Gilded Age. Houghton Mifflin Harcourt. pp. 44-45. ISBN 0618082409.
- ^ Liddell, Henry George och Robert Scott (1980). A Greek-English Lexicon (förkortad utgåva). United Kingdom: Oxford University Press. ISBN 0-19-910207-4.
- ^ Naomi Lubick, Investigating the Antarctic, Geotimes, 2005.
- ^ Erwin DH (1993). The great Paleozoic crisis; Life and death in the Permian. Columbia University Press. ISBN 0231074670.
- ^ Cite error: Invalid
<ref>tag;no text was provided for refs namedconsolation - ^ G.H Groenewald (1991). Burrow casts from the Lystrosaurus-Procolophon Assemblage-zone, Karoo Sequence, South Africa, African Protected Area Conservation and Science 34 (1): 13-22.
- ^ GM King, MA Cluver (1990). Den vattenlevande Lystrosaurus: en alternativ livsstil. Historical Biology: An International Journal of Paleobiology, 1029-2381, 4 (3):323-341.
- ^ Cite error: Ogiltig
<ref>-tagg; ingen text har tillhandahållits för referenser som heterBenton2006WhenLifeNearlyDied