|
Quick facts for kids
Edaphosaurus Temporal range: Górny karbon do dolnego permu, 300-280 mya
|
|
|---|---|
 |
|
| Edaphosaurus at AMNH | |
| Klasyfikacja naukowa | |
| Kingdom: | |
| Phylum: | |
| Subphylum: | |
| Class: | |
| Order: | |
| Family: |
Edaphosauridae
|
| Genus: |
Edaphosaurus
Cope, 1882
|
Edaphosaurus był pelykozaurem (wczesne Synapsida), który żył w późnym karbonie i wczesnym permie. Był roślinożernym krewnym dobrze znanego Dimetrodona. Oba miały duży, termoregulacyjny żagiel na grzbiecie. Skamieniałości Edaphosaurus zostały znalezione w Europie i Ameryce Północnej.
Gatunek Edaphosaurus mierzył od 0,5 metra (1,6 stopy) do prawie 3,5 metra (11,5 stopy) długości i ważył ponad 300 kilogramów (660 lb). Był nieco wyższy i bardziej krępy niż Dimetrodon, ale podobnie jak jego krewni pelykozaury miał małą głowę i długi ogon.
Opis i paleobiologia

Gatunki Edaphosaurus mierzyły od 0,5 metra (1,6 stopy) do prawie 3,5 metra (11,5 stopy) długości i ważyły ponad 300 kilogramów (660 lb). W związku z małą głową, kręgi szyjne są zredukowanej długości, podczas gdy kręgi grzbietowe są masywne, ogon jest głęboki, kończyny krótkie i mocne, a żebra tworzą szeroką klatkę piersiową. Jak większość roślinożerców, Edaphosaurus miałby pojemne jelito i symbiotyczne bakterie pomagające w rozkładzie celulozy i innych niestrawnych materiałów roślinnych. Podobnie jak jego bardziej znany krewny Dimetrodon, Edaphosaurus miał przypominającą żagiel płetwę, która była podtrzymywana przez kości kręgosłupa. Edaphosaurus różni się od Dimetrodona posiadaniem poprzecznych prętów na kolcach, które podtrzymywały jego płetwę.
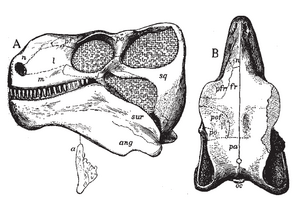
Czaszka
Głowa Edaphosaurus była krótka, stosunkowo szeroka, trójkątna w zarysie i wyjątkowo mała w porównaniu do rozmiarów ciała. Głęboka dolna szczęka prawdopodobnie miała potężne mięśnie, a zęby brzeżne wzdłuż przodu i boków szczęk miały ząbkowane końcówki, co pomagało Edaphosaurus w chwytaniu kawałków twardych roślin lądowych. Tylne części dachu pyska i wnętrze dolnej szczęki zawierały gęste baterie zębów podobnych do kołków, tworzących szeroką powierzchnię miażdżącą i ścierającą z każdej strony powyżej i poniżej. Jego ruchy szczęk były propalinalne (od przodu do tyłu). Wczesne opisy sugerowały, że Edaphosaurus żywił się bezkręgowcami, takimi jak mięczaki, które mógł miażdżyć swoimi płytkami zębowymi. Jednak obecnie paleontolodzy uważają, że Edaphosaurus jadł rośliny, choć ścieranie się zębów między górnymi i dolnymi płytkami zębowymi wskazuje na „ograniczone przetwarzanie pokarmu” w porównaniu z innymi wczesnymi roślinożercami, takimi jak Diadectes, duży nieamniotyczny reptiliomorf (Diadectidae), który żył w tym samym czasie. Wczesnym członkom Edaphosauridae, takim jak Ianthasaurus brakowało płytek zębowych i jedli owady.
Żagiel

Żagiel wzdłuż grzbietu Edaphosaurus był podtrzymywany przez ogromnie wydłużone kręgosłupy nerwowe od szyi do lędźwi, połączone za życia tkanką. W porównaniu z żaglem Dimetrodona, kręgosłupy kręgowe są krótsze i cięższe oraz posiadają liczne małe poprzeczki. Edaphosaurus i inni przedstawiciele Edaphosauridae wykształcili wysokie żagle grzbietowe niezależnie od żyjących w tym samym czasie żaglastych przedstawicieli Sphenacodontidae, takich jak Dimetrodon i Secodontosaurus, co jest niezwykłym przykładem ewolucji równoległej. Funkcja(e) żagla u obu grup jest wciąż przedmiotem dyskusji. Naukowcy sugerują, że takie żagle mogły zapewniać kamuflaż, napędzane wiatrem żeglowanie po wodzie, kotwiczenie dla dodatkowego wsparcia mięśni i usztywnienia kręgosłupa, ochronę przed atakami drapieżników, miejsca magazynowania tłuszczu, powierzchnie regulujące temperaturę ciała, czy też popisy seksualne i rozpoznawanie gatunków. Wysokość żagla, krzywizna grzbietów i kształt poprzeczek są odmienne u każdego z opisanych gatunków Edaphosaurus i wykazują tendencję do większych i bardziej rozbudowanych (ale mniej licznych) procesów projekcyjnych na przestrzeni czasu. Romer i Price zasugerowali, że występy na grzbietach Edaphosaurus mogły być osadzone w tkance pod skórą i mogły służyć do magazynowania pokarmu lub tłuszczu, podobnie jak garb u wielbłąda. Bennett twierdził, że kostne występy na kolcach Edaphosaurus były odsłonięte i mogły tworzyć zawirowania powietrza dla bardziej efektywnego chłodzenia nad powierzchnią żagla w celu regulacji temperatury ciała. Ostatnie badania, w których zbadano mikroskopową strukturę kostną wysokich neuralnych kolców u edafozaurów, wzbudziły wątpliwości co do termoregulacyjnej roli żagla i sugerują, że funkcja wyświetlania jest bardziej prawdopodobna.
Gatunki
| Gatunki | Autorytet | Lokalizacja | Status | Synonimy | Obrazy |
|---|---|---|---|---|---|
| Edaphosaurus boanerges | Romer & Cena, 1940 | Texas | Valid |  |
|
| Edaphosaurus colohistion | Berman, 1979 | West Virginia | Valid | ||
| Edaphosaurus cruciger | Cope, 1878 | Teksas i Oklahoma | Prawdziwy |  |
|
| Edaphosaurus microdus | Cope, 1884 | Texas | Walid | Naosaurus microdus | |
| Edaphosaurus novomexicanus | Williston & Case, 1913 | Nowy Meksyk | Walid |  |
|
| Edaphosaurus pogonias | Cope, 1882 | Texas | Valid |  |
|
| Edaphosaurus raymondi | Case, 1908 | Texas | Valid |
Odkrycie i klasyfikacja

Edward Drinker Cope nazwał i opisał Edaphosaurus („jaszczur chodnikowy”) w 1882 roku na podstawie zmiażdżonej czaszki i lewej dolnej szczęki z Texas Red Beds. Zwrócił on szczególną uwagę na „gęste uzębienie” zarówno na górnej, jak i dolnej szczęce, a w tabeli w opisie użył terminu „chodnik zębowy”. Nazwa gatunkowa pogonias oznacza po grecku „brodaty”, odnosząc się do powiększonego, nachylonego do wewnątrz podbródka na dolnej szczęce. Cope zaklasyfikował Edaphosaurus jako członka swojej Pelycosauria i stworzył nową rodzinę Edaphosauridae. Typowy materiał nie zawierał żadnych elementów szkieletu pozaczaszkowego poza kręgiem osiowym, a Cope nie wiedział o dużym żaglu tego zwierzęcia, który był wówczas znany tylko u Dimetrodon.
W 1886 roku Cope utworzył nowy rodzaj Naosaurus „jaszczur okrętowy” (od greckiego naos „statek”) dla szczątków szkieletowych podobnych do tych u długonosego Dimetrodon, ale z charakterystycznymi „poprzecznymi wyrostkami lub odgałęzieniami, które przypominają ramiona masztu okrętowego”. Spekulował, że „ramiona były połączone błonami z kręgosłupem nerwowym lub masztem, służąc zwierzęciu jako żagiel, z którym poruszało się po wodach permskich jezior”. Rozpoznał trzy gatunki: Naosaurus claviger „club-bearer” (ze względu na występy na kolcach; obecnie uważany za synonim Edaphosaurus pogonias); Naosaurus cruciger „cross-bearer” (ze względu na występy na kolcach; po raz pierwszy opisany przez Cope’a jako Dimetrodon cruciger w 1878 roku; obecnie Edaphosaurus cruciger, największy pod względem rozmiarów gatunek); oraz Naosaurus microdus „small tooth” (po raz pierwszy opisany jako Edaphosaurus microdus w 1884 roku). Cope zauważył niekompletny materiał czaszki znaleziony wraz z okazami N. claviger i N. microdus, ale uważał, że Naosaurus jest odrębny od Edaphosaurus. Później uznał, że Naosaurus musiał mieć dużą mięsożerną czaszkę podobną do Dimetrodon, choć nie miał na to bezpośredniego dowodu kopalnego.
W 1907 roku amerykański paleontolog Ermine Cowles Case zasugerował, że czaszka Edaphosaurus może należeć do szkieletów zwanych Naosaurus, na podstawie okazu znalezionego w 1906 roku, który wydawał się kojarzyć elementy obu. W 1913 roku Samuel Wendell Williston i Case opisali nowy gatunek Edaphosaurus novomexicanus na podstawie dość kompletnego okazu odkopanego w Nowym Meksyku w 1910 roku, w którym znaleziono żaglowaty szkielet typu Naosaurus z małą czaszką typu Edaphosaurus. Starsza nazwa rodzajowa Edaphosaurus Cope, 1882 stała się obowiązującą.
W 1940 roku paleontolodzy Alfred Sherwood Romer i Llewellyn Ivor Price nazwali nowy gatunek Edaphosaurus boanerges („grzmiący orator”) – ironiczne odniesienie do wyjątkowo małych rozmiarów holotypowej dolnej szczęki na złożonym szkielecie pierwotnie zamontowanym w Museum of Comparative Zoology (Uniwersytet Harvarda) z odrestaurowaną głową opartą na większym gatunku Edaphosaurus cruciger.
W 1979 roku paleontolog David Berman utworzył Edaphosaurus colohistion („stunted sail”) dla wczesnego gatunku ze stosunkowo małym żaglem, na podstawie skamieniałości z Zachodniej Wirginii.
Inne proponowane gatunki Edaphosaurus zostały oparte na bardziej fragmentarycznym materiale, który nie może być rygorystycznie zdiagnozowany do poziomu rodzaju/gatunku, ale który mimo to może reprezentować edafozauridy.
W kulturze popularnej
Amerykański paleoartysta Charles R. Knight zrekonstruował Edaphosaurus (jako „Naosaurus”) z czaszką Dimetrodona, którą Cope wcześniej błędnie zaliczył do tego rodzaju. Obraz ten powstał na zamówienie Amerykańskiego Muzeum Historii Naturalnej w 1897 roku i został przedrukowany do nekrologu Cope’a w listopadowym numerze The Century Magazine z 1898 roku. Knight stworzył później dokładniejszą, poprawioną wersję obrazu, która zmieniła „Naosaurus” w Dimetrodon.
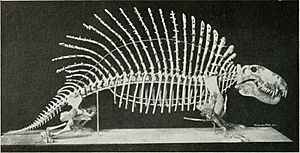
W 1907 roku szkielet „Naosaurus claviger” (synonim Edaphosaurus pogonias) został zamontowany w Amerykańskim Muzeum Historii Naturalnej. Szkielet ten był kompozytem, w skład którego wchodziła czaszka Dimetrodona, jak określił ten gatunek E.D. Cope. Ponieważ „Naosaurus” był uważany za bliskiego krewnego Dimetrodona, a nie Edaphosaurus, do tego złożonego okazu zamontowano także smukłe kończyny należące prawdopodobnie do Dimetrodon dollovianus, zamiast prawidłowych, bardziej krępych. W latach 40. XX wieku ten zamontowany szkielet został zaktualizowany, by odzwierciedlać bardziej aktualną wiedzę na temat Edaphosaurus, z czaszką opartą na tej z E. cruciger.
Artysta Rudolph Zallinger przedstawił Edaphosaurus obok Dimetrodon i Sphenacodon, by reprezentować okres permski w swoim słynnym muralu The Age of Reptiles (1943-1947) w Yale Peabody Museum. Mural został odtworzony z mniejszej wersji obrazu w serii The World We Live In publikowanej w magazynie Life w latach 1952-1954. Seria czasopism została zredagowana w popularną książkę w 1955 roku.
Edaphosaurus pojawia się w odcinku 2 filmu dokumentalnego BBC Walking with Monsters. Pewne stado zostało zaatakowane przez samicę Dimetrodona.